teslabel l belgique
un consensus scientifique croissant concernant les interactions entre biologie cellulaire et moléculaire et les champs électromagnétiques de l’environnement
par
w. ross adey
- Veterans Affairs Medical Center & University School of Medicine, Loma Linda, California, 92357 U.S.A.
- Président du Conseil National Américain de Protection contre les Radiations (n.c.r.p.)
- Mandaté par Environmental Protection Agency (e.p.a.)
In Transmissions électromagnétiques : les dernières évidences scientifiques , nuisances potentielles et stratégies en vue de réduire le risque. Symposium de la Communauté Européenne, Londres, 27 octobre 1994.
1. introduction
Au cours de ces quinze dernières années, des études épidémiologiques ont soulevé des interrogations concernant les risques liés aux expositions aux champs électromagnétiques associés aux réseaux de transport, de distribution d’électricité et à l’usage de l’énergie électrique.
Depuis le début, il apparaît clairement que les composantes en termes d’énergie de ces champs en relation avec les tissus sont extrêmement faibles, tellement faibles, en fait, que la crédibilité à accorder aux études épidémiologiques eu égard à de telles sensibilités, requiert des concepts fondamentalement nouveaux en matière des processus énergétiques de la matière vivante. C’est à ce point que de nouvelles études de laboratoire seront essentielles.
D’autre part, de nouvelles études épidémiologiques semblent ne devoir apporter aucun progrès majeur jusqu’à ce qu’une dose mesurable applicable aux tissus puisse être établie. Des études de laboratoire sont maintenant focalisées sur la physique des processus au niveau des atomes, lesquels processus se déroulent dans le cadre des biomolécules.
Au cours des dix dernières années, des développements mettant en jeu des collaborations remarquables entre les sciences physiques et les sciences biologiques ont réuni ces disciplines apparemment distinctes en un nouveau domaine de la Science. Les recherches sur les états fondamentaux de la matière qui sont désignés par les états de non équilibre et par l’électro- dynamique non linéaire ont suivi leur route main dans la main avec une nouvelle vision intégrant ces concepts situés aux confins de la biologie cellulaire et moléculaire. Par conséquent, il existe maintenant un nouveau projet de définition de la matière vivante en ces termes de physique.
Au delà de la chimie des molécules qui réalise la construction spécifique des tissus vivants, nous devons maintenant discerner une nouvelle frontière peut-être plus difficile à comprendre, dans l’organisation biologique. Ce concept est basé sur les processus physiques au niveau atomique, plutôt que sur les réactions entre les biomolécules. Nous allons voir que ces processus physiques peuvent efficacement réguler les processus des réactions biochimiques.
Notre propos consistera à décrire quelques unes des étapes principales ayant conduit à cette vaste synthèse. Celle-ci décrit même à des niveaux plus ténus, une séquence hiérarchisée d’événements suscités par les interactions moléculaires avec les champs électromagnétiques ambiants. Nous apportons ici notre contribution à la recherche scientifique en faisant référence aux publications les plus expressives confrontées avec les industries productrices d’électricité, aujourd’hui et dans le futur.
2. les basses fréquences électromagnétiques naturelles : modifications attribuables à l’usage de l’énergie électrique
Toute vie sur terre s’est développée dans un environnement de champs électromagnétiques naturels à basses fréquences. Celles-ci ont une source terrestre ou extraterrestre. L’activité orageuse dans les zones équatoriales d’Amérique du Sud ou d’Afrique Centrale produit des champs à extrêmement basses fréquences (E.L.F.) qui se propagent sur toute la planète entre la surface de la terre et l’ionosphère. Cette activité de champs à extrêmement basses fréquences présente une série de pics compris dans une bande s’étalant de 8 à 32 Hertz. Ces pics sont connus sous les termes de résonances Schumann (1957). Leur composante électrique a une intensité voisine de 0,01 Volt/m (10 millivolts/m) dans l’air; leur composante magnétique atteint des intensités de l’ordre de 1 à 10 nanotesla (0,01 à 0,1 mG), ce qui est bien inférieur à l’intensité du champ magnétique statique (continu) située aux alentours de 50.000 nanotesla (500 milligauss).
L’utilisation de plus en plus importante de l’énergie électrique au cours de ce siècle a modifié de manière brutale cet environnement électromagnétique naturel dans le contexte urbain. Les expositions aux champs de ces sources d’énergie sont aujourd’hui, dans nos sociétés civilisées, beaucoup plus intenses que celles dues aux champs naturels. Typiquement, dans les habitations américaines, les niveaux de champs magnétiques alternatifs ambiants peuvent atteindre un ordre de grandeur de 0,3 à 3 milligauss (0,03 à 0,3 microtesla). Ces niveaux peuvent indiquer des valeurs beaucoup plus élevées au voisinage de machines à laver, de réfrigérateurs, de conditionneurs d’air. Les rasoirs électriques, les sèche-cheveux, les couvertures chauffantes produisent tous des champs localisés d’intensité beaucoup plus élevée que celle des niveaux de champs domestiques ambiants. En effet, ils peuvent dépasser les niveaux de champs ambiants produits par les lignes de transport et par les systèmes de distribution d’électricité, mais il faut souligner que ces expositions aux appareils domestiques peuvent n’avoir lieu que sur une très courte période par jour et l’intensité diminue rapidement à une courte distance de ces appareils, tandis que dans des habitations proches de grandes lignes à haute tension, l’exposition s’étale sur de longues durées par jour et typiquement sur plusieurs années.
3. le dilemme biologique et biophysique dans les tissus réputés sensibles à ces niveaux de champs électromagnétiques environnementaux à extrêmement basses fréquences (e.l.f.)
Selon le point de vue des physiciens, certains champs électromagnétiques extrêmement faibles, descriptibles en termes d’énergies de photons, sont nécessaires pour initier les réactions entre systèmes biomoléculaires. Lorsqu’une radiation électromagnétique est décrite en fonction de ces particules photoniques, et lorsque sa longueur d’onde est plus grande que celle correspondant à la région ultraviolette du spectre (350 nanomètres), l’énergie photonique est inférieure à 12 eV (electron-Volt) et est insuffisante pour provoquer l’ionisation des molécules, c’est à dire leur rupture avec formation de radicaux libres. La longueur d’onde associée à la fréquence 50 Hz est 6.000 km.
Pour les biologistes, l’édifice fonctionnel de la matière vivante se limite à l’échange de chaleur et le moteur thermique reste le modèle dans les séquences des processus animés. Nous sommes maintenant confrontés avec un domaine de plus en plus large où l’échange thermique des tissus n’est pas la clé de la réponse. Nous allons discuter de cette évidence en détails. Bien qu’il existe toujours dans certains points de vue des physiciens une opinion tenace selon laquelle les champs électromagnétiques non ionisants seraient incapables d’induire des effets biologiques autres que des effets thermiques, (Foster and Guy, 1986 ; Foster and Pickard, 1987), l’évidence du point de vue opposé acquiert progressivement un consensus international (Adey, 1990, 1993 ; Grundler et al., 1992 ; Terada et al., 1994).
Pour les physiciens et les biologistes, il existe un écueil central, étant donné que ces sensibilités impliqueraient des stimuli situés en deçà du niveau d’énergie intrinsèque des collisions atomiques thermiques (kT), lequel a été considéré comme un seuil pour tous les stimuli biologiques. La base physique de cette sensibilité reste incomplète, mais comme nous allons le voir, des évidences théoriques et expérimentales indiquent maintenant le rôle des radicaux libres, formés très brièvement au cours de périodes de quelques nanosecondes, dans le déroulement de toutes les réactions chimiques. Ils répondent à un mécanisme situé à un niveau microscopique, qui transcende cette barrière de l’énergie thermique dans la première étape transductrice de l’interaction entre champs et tissus. La sensibilité des radicaux libres aux champs magnétiques peut être étendue à toutes les voies jusqu’à des champs d’énergie proches de zéro (Grundler et al., 1992 ; Mc Lauchlan, 1992).
Les systèmes biomoléculaires présentent de hauts niveaux de coopération comme base de réponse aux champs athermiques à extrêmement basses fréquences (E.L.F.), et aux champs de fréquences radio, modulées en amplitude à des fréquences E.L.F. (Adey, 1990, 1993 pour les revues). En bref, cette coopération se révèle dépendre en partie d’états cohérents dans des populations de charges électriques sur des chaînons de protéines. Il existe des études indépendantes concernant les effets biologiques des champs magnétiques à extrêmement basses fréquences (E.L.F.) qui montrent une dépendance spécifique vis-à-vis de combinaisons de champs magnétiques continus et alternatifs.
Des études théoriques et expérimentales discutent des résonances à basses fréquences pour des combinaisons de champs alternatifs et continus variables en intensité (Liboff, 1992) ainsi que des phénomènes fenêtre pour des sensibilités liées aux amplitudes de champs alternatifs. Des modèles théoriques applicables à ces phénomènes montrent l’existence d’un problème commun d’actions conjointes du bruit de fond thermique et électrique inhérents aux cellules vivantes, à des températures normales. Le problème exprimé par ces modèles physiques est en relation avec des échelles de temps de l’ordre de la nanoseconde (10-9 seconde) des collisions thermiques, qui pourraient tendre à interrompre les temps de cohérence de l’ordre de la milliseconde, essentiels pour obtenir des interactions résonantes avec un champ appliqué à des fréquences situées entre 1 et 100 Hertz.
4. eléments des tissus dans la détection des champs électromagnétiques - le rôle des membranes cellulaires
Au delà de ces événements physiques entrant en jeu dans la première détection des champs électromagnétiques environnementaux dans les tissus, un consensus général semble s’imposer; il consiste à admettre que le site d’action des champs est la membrane cellulaire.
Des chaînons de protéines sont stratégiquement situés à la surface des cellules dans les tissus. Ils y agissent comme détecteurs de messages électriques et chimiques arrivant à la surface des cellules, les traduisent et les transmettent à l‘intérieur des cellules. La base structurelle de ce couplage de transduction par ces chaînons protéiques est bien connue. Grâce à eux, les cellules accomplissent un triple rôle: la détection des signaux, l’amplification des signaux, la transmission des signaux à l’intérieur des cellules.
Les ions calcium (Ca++) jouent un rôle essentiel à chaque étape de cette signalisation transmembranaire et ont été utilisés comme marqueurs des interactions de champs électromagnétiques avec une quantité de tissus et de cultures cellulaires. Les études concernant la fuite du calcium (efflux calcique), d’abord dans les tissus cérébraux, ont révélé des sensibilités aux champs électromagnétiques à extrêmement basses fréquences (E.L.F.) et aux champs de fréquences-radio modulées en E.L.F. (Bawin et al., 1975, 1976 ; Blackman et al., 1979, 1985 a, b, 1994 ; Dutta et al., 1984 ; Lin-Liu and Adey, 1982). Dans beaucoup de cas, ces réponses obéissaient à un effet fenêtre en ce qui concerne l’amplitude du champ ou la fréquence du champ E.L.F. ou la modulation E.L.F. de la fréquence-radio, ou les deux à la fois. Des études plus récentes concernant les fuites de calcium ont suscité un consensus plus étendu sur le rôle clé du calcium.
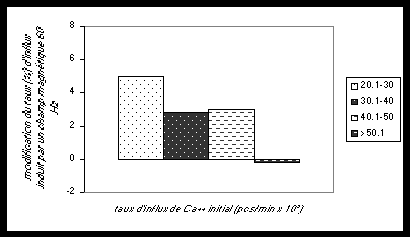 |
Fig. 1 . Effets de l’exposition
de cellules leucémiques T Jurkat humaines pendant deux minutes à des champs
magnétiques alternatifs 60Hz. Le taux de capture de Ca++ (Mn++) (influx) est
modifié. Les résultats sont exprimés en moyennes de sensibilités électromagnétiques
par des comptages photoniques (p. c. sec. / min). La signification statistique
a été vérifiée par les tests pairés et anova.
Les mesures de capture de
calcium par les lymphocytes chimiquement activés grâce à un mitogène et ensuite
exposés à un champs magnétique 3Hz 2 mt ont montré qu’avec un faible
taux de capture (20.000 à 30.000 comptages / min.), on pouvait observer une
diminution de 5% de capture. Par contre avec de forts taux de capture (au
dessus de 50.000 comptages/min.), les lymphocytes ne répondaient pas au champs.
La sensibilité au champ magnétique a été préalablement déterminée par les
conditions initiales des lymphocytes.
Ces études ont été focalisées sur les cellules du système immunitaire (Lyle et al., 1991 ; Walleczek, 1992 ; Walleczek and Budinger, 1992). Dans deux études indépendantes, réalisées avec des lymphocytes mitogènes-activés, l’exposition à un champ magnétique pulsé de 3 Hz de l’ordre de grandeur du millitesla (10.000 mG) a fortement réduit la capture d’ions calcium (Conti et al.,1985 a & b ; Walleczek, 1992). Par contre, des stimulations à 60 Hz dans les mêmes conditions ont fortement augmenté l’absorption de calcium (Walleczek and Liburdy, 1990).
Deux autres études indépendantes, utilisant des champs magnétiques pulsés de 3 Hz ont révélé une réduction de 55 à 60 % de la capture de thymidine par des lymphocytes mitogènes-activés (Conti et al., 1985 a & b ; Mooney et al., 1986). Ces observations mettent en exergue l’importance de l’état d’activation des cellules au début de l’exposition aux champs électromagnétiques, étant donné que les cellules non encore stimulées par les mitogènes ne répondent pas aux champs électromagnétiques.
5. signalisation interne aux membranes cellulaires : les champs électromagnétiques et une cascade d’enzymes
La modulation de la liaison entre les ions calcium et la surface cellulaire par les champs électromagnétiques est à mettre en relation avec un important système de coopération, car les modifications induites per les champs dans la fuite du calcium des tissus sont bien plus importantes que celles que l’on aurait pu pronostiquer en fonction de l’énergie appliquée. Elle est donc en rapport avec une très forte amplification du signal de déclenchement. Ce signal amplifié est transmis le long des chaînons des protéines réceptrices à l’intérieur des cellules, où il active des enzymes situées sur ou à proximité des membranes cellulaires. Ces enzymes sont des molécules de protéines qui fonctionnent comme des catalyseurs, servant d’intermédiaires aux réactions chimiques à la température des tissus. Ces réactions chimiques, en l’absence de ces catalyseurs, n’auraient lieu qu’à des niveaux d’énergie beaucoup plus élevés (p.ex. température et pression). Ces enzymes subsistent après réactions et peuvent ainsi participer à un même processus de manière répétitive.
L’amplification du signal transmembranaire se prolonge lorsque ce signal atteint l’intérieur de la cellule. Beaucoup de signaux transmis à travers la membrane par le chaînon récepteur entrent d’abord en interaction avec une classe de protéines connues sous les termes protéines G, ainsi dénommées parce qu’elles sont liées à la guanosine triphosphate (GTP). Au cours d‘une séquence ultérieure, la guanosine triphosphate peut stimuler ou inhiber la puissante enzyme métabolique adénylate-cyclase (ATP). Un chaînon protéique récepteur peut activer beaucoup de molécules de protéines G (Luben, 1990), induisant ainsi d’autres amplifications du signal. Les champs magnétiques désensibilisent les récepteurs liés aux protéines G. Leur importance dans les transmissions de signaux à l’intérieur des cellules a été reconnue par Gilman et Rodbell, en 1994, lauréats du prix Nobel.
A partir de leur entrée dans la cellule, les signaux transmembranaires activent une cascade d’enzymes. Dans le but que nous poursuivons ici, nous devons limiter nos considérations à une série d’enzymes identifiées comme ayant des effets d’interactions avec les champs électromagnétiques appliqués. Cette cascade implique des enzymes métaboliques, des enzymes messagères, des enzymes de croissance et des enzymes de régulation.
a. L’activité métabolique cellulaire est nourrie par l’adénosine triphosphate (ATP). La défaillance de l’ATP implique l’enzyme en relation avec la membrane cellulaire: l’adénylate- cyclase. Une évidence quant à la sensibilité de l’adénylate-cyclase aux champs électromagnétiques repose sur des études concernant son rôle dans la consolidation des fractures (Luben, 1989, 1990 ; Luben and Cain, 1984 ; Luben et al., 1982). Des signaux provenant d’hormones liées aux récepteurs de la surface cellulaire activent l’adénylate-cyclase. Cette activation est fortement modulée par une quantité de champs magnétiques à basses fréquences.
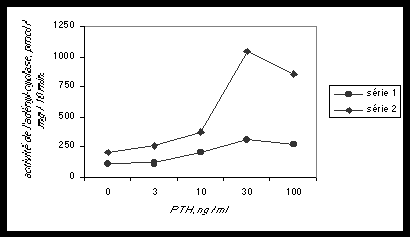
PTH . parathyroïde hormone ..sans champs EM -Naf = 1144 +/- 231
PTH . parathyroïde hormone . . avec champ E.M. 72 Hz, 90 heures – NaF = 1379 +/- 306
Fig.2 l L’activité de l’adényl-cyclase dans les cellules osseuses est stimulée par l’hormone parathyroïde (PTH). La courbe supérieure représente cette stimulation (qui aboutit à la destruction du tissu osseux) en fonction de la concentration en PTH, en l’absence de champ magnétique pulsé. La courbe inférieure montre une nette inhibition de cette stimulation au niveau de l’adényl-cyclase par une exposition de 90 heures à un champ magnétique pulsé de 72 Hz. On voit donc que les champs magnétiques pulsés à la fréquence de 72 Hz contribuent à reconstruire le tissu osseux en inhibant l’activité de l’adényl-cyclase. Ce traitement de fracture est utilisé aux USA depuis 1972.En enlevant des groupes phosphates, l’adénylate-cyclase transforme l’ATP en 3',5'-adénosine monophosphate cyclique (cAMP), avec libération d’une énergie calorifique considérable. A son tour, le cAMP prolonge la cascade enzymatique en transférant des groupes phosphates à une famille d’enzymes-messagères, les protéines-kinases.
b. Les enzymes-messagères protéines-kinases, activées par ces signaux en relation avec la membrane cellulaire, peuvent se disperser largement à l’intérieur de la cellule, diffusant des signaux vers beaucoup de structures, y compris à l’intention du noyau cellulaire. Leur activité est modulée par les champs électromagnétiques à extrêmement basses fréquences et par des champs de fréquences-radio modulées par des E.L.F. (Byus et al., 1984 ; Luben, 1994).
c. La croissance des cellules et la synthèse de l’ADN impliquent l’enzyme ornithine-décarboxylase (ODC) dans les étapes-clés de la régulation. L’ornithine-décarboxylase synthétise les polyamines (putrescine, spermidine, spermine) à partir de l’acide aminé ornithine. Les polyamines semblables à des serpentins, ont le rapport charge/masse le plus élevé de toutes les biomolécules. L’activité de l’ornithine-décarboxylase est modulée par les champs électriques et magnétiques à extrêmement basses fréquences (E.L.F.) (Byus et al., 1987 ; Cain et al., 1993) et par les champs de fréquences-radio modulées par des extrêmement basses fréquences (Byus et al., 1988 ; Litovitz et al., 1993).
Dans les noyaux cellulaires, les polyamines sont essentielles dans la synthèse de l’ADN. Des niveaux élevés de polyamines à l’intérieur des cellules déclenchent également la transcription des proto-oncogènes c-myc et c-fos par c-ras. Ainsi, les polyamines peuvent participer à une cascade d’événements conduisant à la communication entre des produits oncogènes liés à la membrane cellulaire et à des produits oncogènes du noyau (Tabib and Bachrach, 1994).
Les polyamines sont également exportées vers la surface cellulaire (Byus et al., 1994) où elles exercent un rôle plutôt séparé (Mc Bain and Mayer, Revue, 1994) soit en stimulant, soit en inhibant le récepteur spécifique (récepteur NMDA) de l’acide aminé acide L-glutamique qui contribue à l’excitation des transmissions synaptiques à des sites situés dans le cerveau et au niveau de la moelle épinière. Les actions stimulatrices ou inhibitrices des polyamines dépendent du niveau de potentiel de la membrane cellulaire et de la molécule spécifique de polyamine. Ainsi, on voit apparaître un modèle expérimental, dans lequel les champs électromagnétiques à des niveaux athermiques, donc inférieurs aux niveaux de dégagement de chaleur, sont d’abord perçus par les récepteurs de la surface cellulaire, amplifiés puis signalés à l’intérieur de la cellule, et ensuite, grâce à une cascade d’enzymes, ils déterminent la production de modulateurs chimiques, parmi lesquels quelques uns sont renvoyés vers la surface cellulaire pour y répéter le premier cycle d’activité. Des signaux provenant de cette cascade initient également des transcriptions de gènes dans l’acide désoxyribonucléique (ADN) (Phillips, 1993).
6. signaux externes traversant la membrane cellulaire; champs électromagnétiques comme modulateurs de la communication de cellule à cellule dans la régulation de la croissance cellulaire
Les cellules dans les tissus normaux, peuvent être considérées comme une société organisée, chuchotant entre elles dans un langage discret et privé. Elles communiquent avec leurs voisines via des stimuli chimiques et via des champs électriques bien plus faibles que ceux dus au potentiel de la barrière électrique établie par la membrane. Des jonctions par des trous, formés de plaques spécialisées de protéines placées entre les membranes de cellules voisines, fournissent les couplages électriques et chimiques entre elles (Löwenstein, 1981).
Elles sont perforées de nombreux tubuli très fins (connexons) formant des passages à travers lesquels des molécules synthétisées dans une cellule atteignent le cytoplasme de cellules voisines. Cette coopération métabolique est essentielle dans la régulation de la croissance normale (Pitts and Finbow, 1986).
Par contre, le cancer peut être considéré comme une rébellion dans une société organisée de cellules. Les cellules cancéreuses ne tiennent pas compte de leurs voisines et croissent de façon autonome, sur les cellules normales environnantes. Etant donné que la communication intercellulaire joue un rôle important dans le maintien de la société organisée, elle doit être perturbée au cours du processus de carcinogénèse (Yamasaki, 1987, 1990). Des interruptions de communication via ces jonctions par trous peuvent conduire à des dysrégulations de croissance cellulaire (Adey, 1992 pour la revue). Ce processus peut être inversé dans des cultures de cellules cancéreuses, si on les remet en contact avec des cellules saines (Newmark, 1987). Des oncogènes viraux spécifiques causent des effets différentiels dans la communication de cellule à cellule, et arrivent ainsi à supprimer la croissance des cellules cancéreuses non régulée par des cellules normales, lorsque les deux types de cellules sont cultivées ensemble (Bignami et al., 1988). L’introduction d’un oncogène H-ras-1 activé dans des cellules épithéliales réduit significativement la communication via les jonctions par trous. Il est possible que cette réduction soit due à une décroissance de la phosphorylation dépendant du c-AMP (voir section 5a concernant la protéine principale de jonction par trous).
Des évidences expérimentales confortent le rôle des champs électromagnétiques dans la modulation de la communication via ces jonctions par trous. Nous allons prendre ceci en considération dans le cadre de modèles couramment acceptés de carcinogénèse et de formation de tumeurs.
7. carcinogénèse en étapes: initiation, promotion et progression
Il existe un consensus consistant à admettre que la formation de tumeurs implique au moins deux étapes; une étape précoce d’initiation et une étape postérieure d’effet de promotion. Un simple agent peut à la fois provoquer les deux événements ou deux ou plusieurs agents distincts peuvent être nécessaires, en agissant ensemble pour accomplir la séquence en question. Le temps séparant l’exposition à un agent initiateur de l’apparition de la maladie (période de latence) est souvent de 20 ans ou plus.
L’initiation implique des lésions au matériel génétique de l’ADN situé dans les noyaux cellulaires, mais les modifications ne sont pas exprimées, c’est à dire qu’une tumeur n’en résultera pas avant qu’un ou plusieurs agents promoteurs n’agissent dans la suite, de façon répétitive. L’initiation peut être un événement isolé, tel l’exposition à des rayons X ou l’irradiation lors d’une explosion nucléaire, ou l’exposition à certains produits chimiques comprenant les dérivés de goudron. Les cellules initiées sont transformées (mutées). Ce sont des cellules cancéreuses, mais elles restent au repos si elles ne sont pas stimulées par un agent promoteur. La promotion résulte de l’action d’agents n’ayant pas ou n’ayant que peu d’activité d’initiation lorsqu’ils sont testés seuls, mais qui exaltent nettement la formation de tumeurs lorsqu’ils sont appliqués répétitivement et par intermittence après une faible dose d’un agent initiateur (Slaga et al., 1978 ; Berridge et al., 1988 ; Nishizuka, 1983). Ils n’agissent pas sur l’ADN du noyau cellulaire et beaucoup d’entre eux sont connus pour avoir le pouvoir de se lier aux récepteurs sur la membrane cellulaire (Weinstein, 1988).
8. carcinogénèse épigénétique (non génotoxique) : évidence de l’action des champs électromagnétiques dans la promotion tumorale
De nouvelles voies de recherche dans la formation de tumeurs reflètent l’identification d’un nombre croissant d’agents promoteurs (promoteurs tumoraux), lesquels se révèlent jouer un rôle causal dans les cancers humains, sans action directe sur le matériel génétique de l’ADN. Pitot et Dragan (1991) ont résumé l’importance de cette approche comme un angle d’attaque essentiel dans la recherche actuelle et future sur le cancer.
Dans le domaine de la génotoxicité, le modèle mentionné ci-dessus, se référant à des étapes multiples prévoit des mécanismes multiples et de multiples stades définis d’initiation, de promotion et de progression. Des transitions entre les stades successifs peuvent être exaltés ou inhibés par divers types d’agents. Le développement d’une tumeur maligne implique des interactions complexes entre des facteurs environnementaux (produits chimiques, radiations ionisantes et non ionisantes, virus) et des facteurs endocriniens (génétiques, hormonaux) (Weinstein, 1988).
Le modèle épigénétique (non génotoxique) est focalisé sur l’action d’agents promoteurs de tumeurs, dont beaucoup agissent lors des interactions primaires avec les récepteurs associés aux membranes cellulaires (voir section 3.). Le promoteur chimique le plus utilisé dans l’étude expérimentale des cancers est l’ester de phorbol TPA. Avec la découverte de l’activation de l’enzyme-messagère liée à la membrane protéine-kinase C (PKC) par le TPA, (Castagna et al., 1982) et grâce à d’autres études ultérieures, le PKC s’est révélé être le récepteur cellulaire principal du TPA (Nishizuka, 1983, 1984). Une série de ponts réunissent maintenant la recherche concernant la promotion des tumeurs, les facteurs de croissance, les signaux de transduction, incluant l’action des champs électromagnétiques et l’action d’oncogènes spécifiques (Phillips et al., 1992, 1993; Adey, revues 1990, 1992).
Des évidences expérimentales à partir d’études sur cellules et sur animaux confortent un modèle d’actions conjuguées entre des promoteurs tumoraux chimiques et les champs électromagnétiques au niveau de la membrane cellulaire.
a. modèle cellulaire de promotion. Lorsqu’on réunit dans des cultures cellulaires (co-cultures), des cellules normales (fibroblastes) et leurs cellules-filles préalablement mutées par de la lumière ultraviolette et se comportant comme des cellules cancéreuses, avec une croissance incontrôlée, le contact avec les cellules parentales inhibe la croissance désordonnée. Le promoteur tumoral TPA rompt cet équilibre de contact, permettant ainsi le retour à la croissance incontrôlée des cellules-filles mutantes et la formation de petites tumeurs (foci) dans le puits de culture.
L’exposition de ce système à un champ magnétique 60 Hz (sinusoïdal, 0,1 mT, 1 heure d’exposition, 4 fois par jour) augmente de 60 % le nombre de petites tumeurs (foci) induites par TPA (p < 0,001).
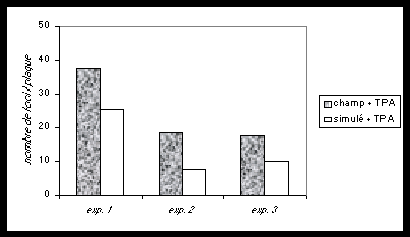
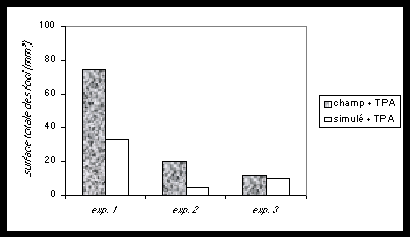
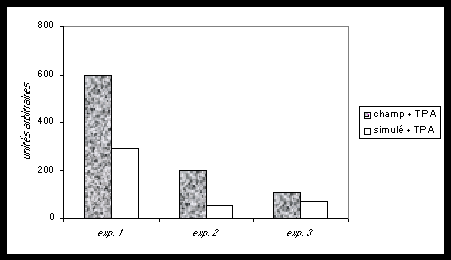
Fig.3 .Cellules normales de fibroblastes et leurs cellules filles mutantes, mises en culture ensemble (co-cultures). Les cellules-mères inhibent la multiplication incontrôlée des cellules mutantes. Cet équilibre est perturbé si un promoteur chimique du cancer (T.P.A.) est ajouté à la culture commune (co-cultures). Les foci (tumeurs en plaques) réapparaissent. Leur nombre, dimensions et densités sont doublés en présence d’un champ magnétique 60Hz de 1 Gauss (d’après Cain et al., 1993).
On constate dans ce cas que ces petites tumeurs ont environ deux fois la dimension et la densité cellulaire des cultures réalisées sans champ magnétique (Cain et al.,1993). Les champs magnétiques seuls n’ont pas d’effet.
Dans ce système, les cellules croissent jusqu’à confluence en 10 à 14 jours, et jusqu’à une monocouche en 28 jours. Ainsi, les expositions intermittentes ont lieu pendant que les cellules croissent en présence de TPA. Ceci suggère que les champs magnétiques 60 Hertz agissent en conjonction avec les promoteurs tumoraux chimiques pour exalter le développement et l’expression des cellules cancéreuses-filles dispersées parmi les cellules-mères. Cette action conjointe des promoteurs chimiques et des champs électromagnétiques est considérée comme une co-promotion.
b. Promotion de cancer sur un modèle de peau de souris. Une étude animale conduite en comparaison avec des sujets-témoins sur l’évolution de cultures cellulaires a utilisé des souris dans une expérimentation initiation-promotion (Mc Lean et al., 1991). Les auteurs ont initié des cancers sur la peau du dos de souris au moyen d’une dose de dimethylbenzanthracène (DMBA) (carcinogène bien connu) située en dessous du seuil de carcinogénicité. Les souris furent alors exposées à un champ magnétique alternatif 60 Hz, à une intensité de 2mT pendant 21 semaines, afin de constater si le champ magnétique pouvait agir comme promoteur tumoral. Aucune tumeur ne s’est développée, ni dans le lot témoin (non exposé), ni dans le lot exposé aux champs magnétiques. Deux groupes supplémentaires furent alors traités une fois par semaine avec le promoteur tumoral TPA. Le délai d’apparition de tumeurs s’est raccourci (mais non de manière statistiquement significative) dans le groupe exposé à la fois aux champs magnétiques et au TPA. Cette étude qui révèle des limite dues au petit nombre d’échantillons, suggère néanmoins une diminution de la surveillance immunitaire par l’activité cellulaire des cellules tueuses naturelles (natural killers = NK). Ces dernières auraient dû dans des circonstances plus favorables prévenir ou retarder la croissance de quelques cellules tumorales.
9. des systèmes cellulaires aux tissus puis aux organes : évidence de l’action des champs électromagnétiques à extrêmement basses fréquences (e.l.f.) sur les réponses immunitaires et sur les mécanismes neuro-endocriniens du cerveau.
La surveillance immunitaire protège le corps contre les infections et les tentacules envahissants du cancer, en distinguant le soi du non soi.
L’immunité cellulaire utilise comme médiateurs les lymphocytes circulant dans le sang et les tissus. Elle se différencie de l’immunité humorale qui utilise comme médiateurs des anticorps circulant dans le sang. L’immunité cellulaire comprend les cellules tueuses naturelles (NK) mentionnées ci-dessus. Les cellules NK sont présentes dans la plupart des organes et sans avoir acquis aucune immunisation antérieure, sont capables de reconnaître, de s’accoler aux cellules tumorales et de les détruire (lyse). Des lymphocytes peuvent également prendre pour cibles des cellules tumorales et les détruire par un contact réel (cytotoxicité allogénique). Dans les études sur cultures cellulaires, cette aptitude à tuer est réduite par des champs électriques alternatifs 60 Hz (Lyle et al., 1988) ainsi que par des champs de fréquences radio modulées par des extrêmement basses fréquences (E.L.F.) (Lyle et al., 1983). Des lymphocytes d’amygdales humaines mis en culture sont également sensibles aux champs de fréquences radio, modulées par des E.L.F. en répondant par une diminution nette mais passagère de l’activité de la protéine-kinase messagère (Byus et al., 1984).
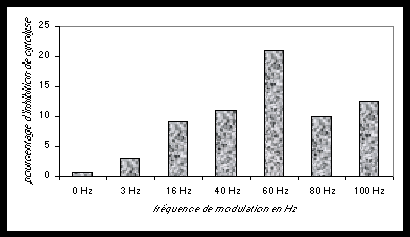
Fig. 4 . Inhibition de la cytotoxicité de lymphocytes T allogéniques par exposition à un champ électromagnétique de 450 MHz (1,5 mW / cm²) dont l’amplitude a été modulée en extrêmement basses fréquences sinusoïdales (E.L.F.) entre 0 et 100 Hz. Cette expérience montre que la sensibilité est sélective (effet fenêtre) en fonction de la fréquence E.L.F. de modulation.
On peut donc conclure que l’aptitude destructrice des lymphocytes vis-à-vis des cellules cancéreuses est diminuée lorsqu’un champ de fréquence radio modulée par des extrêmement basses fréquences est appliqué. La diminution maximale est au-delà de 20% avec une modulation à 60 Hz, la fréquence du réseau électrique américain et une forte composante de modulation des fours à micro-ondes (d’après Lyle et al., 1983).Les sensibilités neuro-endocrines cérébrales vis-à-vis des champs électromagnétiques ont été centrées sur des études concernant la mélatonine, une hormone sécrétée par la glande pinéale (épiphyse). La mélatonine est synthétisée et sécrétée selon un rythme circadien puissant, atteignant un pic nocturne maximal vers 2 heures du matin, chez l’homme et chez les animaux (Reiter, 1986). Le cycle est sensible aux variations du taux d’exposition à la lumière au cours de la nuit et du jour pour différentes espèces. La question de sa variabilité en fonction d‘une modification des champs électromagnétiques ambiants a été l’objet d’études importantes (Semm, 1983 ; Wilson et al., 1981, 1990). Quelques résultats de ces études restent peu clairs selon les espèces et même dans le cadre précis de certaines espèces. Les résultats les plus consistants ont été obtenus par l’exposition de hamsters Djungarian à des champs magnétiques (Yellon, 1994).
Des expositions aiguës de hamsters pendant des jours longs à des champs magnétiques alternatifs 60 Hz (0,1 mT, 15 min), deux heures avant l’extinction des lumières ont supprimé l’augmentation nocturne de mélatonine dans la glande pinéale et dans le sang. Des expositions aigües d’animaux pendant des jours courts ont donné des résultats semblables (Yellon, 1994).
Par contre, des expositions journalières du même type pendant trois semaines n?ont pas indiqué d’effet.
Au delà des rythmes de l’activité diurne, la mélatonine est la clé d’un large éventail de mécanismes de régulation (Reiter, 1992), englobant le système immunitaire, réduisant l’incidence de certains cancers chez les souris et inhibant la croissance de cellules de cancer du sein (Hill and Blask, 1988). Certains chercheurs ont observé que cette activité inhibitrice de la mélatonine sur la croissance du cancer du sein humain est bloquée par des champs magnétiques 60 Hz à un niveau-seuil de 1,2 µT (Lidbury et al.,1994). De plus, des patients porteurs de cancers du sein positifs aux récepteurs d’oestrogènes avaient des niveaux plus bas de mélatonine nocturne dans le plasma (Tamarkin et al., 1982).
10. détection par les tissus de champs électromagnétiques non ionisants à des niveaux athermiques : la recherche de la première étape de transduction dans les mécanismes des radicaux libres.
Nous avons décrit les principales étapes au cours desquelles nous avons découvert une hiérarchie énergétique dont les échelons réalisent des couplages entre l’intérieur de la cellule et les signaux de la surface cellulaire ayant pour origine les champs électromagnétiques à extrêmement basses fréquences (E.L.F.). Notre point de départ n’avait pas pris en considération la première étape tellement importante dans cette séquence: la première détection des champs électromagnétiques, dont l’intensité a souvent un ordre de grandeur inférieur à celui de l’énergie moyenne d’agitation thermique kT dans le substrat biomoléculaire de détection.
Des considérations thermodynamiques concernant l’énergie thermique des tissus n’imposent pas nécessairement un seuil limite dans cette première interaction. Des états non thermiques et l’électrodynamique non linéaire dans les systèmes biologiques offrent un grand nombre de possibilités (Adey, 1981, 1993 ; Adey and Lawrence, 1984 ; Grundler et al., 1992 ; Fröhlich, 1988 ; Kaiser, 1988 ; Walleczek, 1994). Une réponse à cette importante question peut être trouvée dans les interactions des champs électromagnétiques avec les radicaux libres.
Les radicaux libres sont des atomes ou des molécules possédant un ou plusieurs électrons non appariés; c’est cette particularité qui rend les radicaux libres extrêmement réactifs (Walleczek, 1994). Dans les réactions chimiques, les liaisons se rompent puis se reforment. La plupart des réactions chimiques entre des atomes mettent en oeuvre des électrons par paires avec des spins opposés; un électron provenant de chaque partenaire dans l’union. L’électron en rotation (spin) engendre un champ magnétique. Les liaisons chimiques se produisent entre des atomes ayant des spins opposés d’électrons, donc des champs magnétiques de polarisations opposées.
Dans une réaction chimique, la liaison se rompt et chaque partenaire reprend son électron de la liaison, et va à la recherche d’un nouveau partenaire. Chaque partenaire est maintenant libre; il s’agit d’un radical libre très réactif. La reformation d’une liaison exige une rencontre entre deux radicaux ayant des spins électroniques opposés, l’union produisant une paire de singulets. La durée de vie de radicaux libres est typiquement courte, de l’ordre de grandeur de la nanoseconde (10-9 seconde) à la microseconde (10-3 seconde). C’est au cours de cette brève période que des champs magnétiques peuvent altérer la vitesse et le rendement en produits d’une réaction chimique. Etant donné que l’action ne se produit que sur les vitesses (cinétique) des réactions chimiques, ces effets sont appelés effets magnétocinétiques (Steiner and Ulrich, 1989). Ils ne se produisent que dans des états non thermiques de systèmes biomoléculaires, définis comme une insensibilité aux interactions thermiques aléatoires durant la brève période de leur existence (Walleczek, 1994). Ils sont la conséquence d’un saut quantique cohérent qui accompagne la formation d’un radical libre. Mc Lauchlan (1992) a proposé un rôle pour les radicaux libres dans la médiation des interactions moléculaires avec les champs magnétiques 50 Hz et 60 Hz des sources d’énergies électriques. Dans son modèle de très faibles champs magnétiques statiques causent la rupture de triplets et donnent des singulets. A de plus hauts niveaux , situés aux alentours de 8 mT, deux tiers des radicaux ne peuvent pas réagir comme ils le feraient dans un champ plus faible.Un énorme effet d’un faible champ magnétique sur une réaction chimique et l’effet commence à la force la plus faible de champ magnétique appliqué... L’interaction globale a une énergie beaucoup plus faible que celle de l’énergie thermique du système et agit exclusivement par son influence sur la vitesse de réaction. Ceci va à l’encontre de l’intuition des scientifiques!
Bien que les énergies de spin électronique sont conservées au cours des collisions thermiques, leur courte durée de vie en relation avec les fréquences des collisions a soulevé des questions sans réponses à propos des réactions radical-radical. Dans une synthèse mettant en exergue les interactions athermiques entre les champs électromagnétiques et les systèmes cellulaires, Grundler et al. (1992) ont présenté quelques séquences des couplages par transduction des champs électromagnétiques, basées sur des réactions chimiques influencées par des champs électromagnétiques. Ces réactions comprenaient les réactions du cytochrome catalysées par des réactions enzymatiques qui mettent en oeuvre des paires de radicaux et la production de radicaux libres, tels de l’oxygène très réactif et l’oxyde nitrique conduisant à des amplifications hautement coopératives ultérieures. Ils concluent que des champs magnétiques appliqués peuvent être actifs, même à des niveaux proches de zéro. En d’autres termes, un seuil ne peut être établi pour ces systèmes. Il existe une évidence expérimentale nette, confortant le modèle basé sur les radicaux libres couplés avec des transductions de champs électromagnétiques dans les systèmes biologiques.
On peut estimer que plus de 20 enzymes sont susceptibles de s’insérer selon de tels processus dans des couplages radicalaires en vue de convertir des substrats en produits (Harkins and Grissom, 1994). On peut citer comme exemple le fait que des champs magnétiques statiques de l’ordre de grandeur de 0,1 à 0,15 Teslas ralentissent la vitesse de réaction de l’enzyme éthanolamine-ammonia-lyase de 25 à 60 %; ceci s’explique par le fait de modifications induites par les champs magnétiques dans les taux de ruptures entre états singulets et triplets de radicaux. On peut également noter que les enzymes P-450 du cytochrome dépendent de radicaux libres et peuvent être modulées par le P-450 econazole proposé. Cet agent complètement inhibé (même inversé) augmente la capture de calcium dans les lymphocytes T humains et est influencé par un champ magnétique 60 Hz à une intensité de 2 mT(Walleczek et al., 1994).
L’oxyde nitrique (NO) est un radical libre puissant, synthétisé dans beaucoup de tissus. Comme il s’agit d’une molécule gazeuse, il diffuse rapidement bien au delà de la cellule d’origine. C’est un modulateur physiologique important dans le cerveau, la moelle épinière. Il stimule les tissus vasculaires et sert de médiateur dans l’érection pénienne. Il a été impliqué dans les pathologies liées au stress, comprenant les pathologies coronariennes, les maladies de Parkinson et les maladies d’Alzheimer. L’oxyde nitrique est un modulateur de signaux régulièrement récurrents des ondes électriques à rythme lent (E.E.G.) dans les tissus cérébraux. Des champs magnétiques appliqués ( 1 Hz, 0,1 mT) rompent la régularité de ces rythmes d’ondes via un mécanisme dépendant de l’oxyde nitrique NO (Bawin et al.,1994). Des champs de fréquences radio pulsées augmentent la quantité de NO et les niveaux de GMP-cyclique dans le cervelet du rat (Miura et al., 1993).
11. concernant les effets de doses cumulées.
En général, les données épidémiologiques suggèrent une relation entre la durée de l’exposition à des champs électromagnétiques ambiants typiquement mesurés sur plusieurs années et le seuil d’apparition d’un processus pathologique. Il va de soi que ces expositions à long terme sont intermittentes; elles sont mesurées, soit selon une base journalière, soit selon une plus longue échelle de temps. Cependant, l’utilisation d’une moyenne pondérée de temps (Time Weighted Average =T.W.A.) comme mesure d’exposition a une signification très limitée dans la recherche d’une corrélation précise avec le risque de maladie. D’autres mesures de l’environnement électromagnétique comme les niveaux de champs ambiants ne permettent guère d’établir une mesure fiable dose-réponse. Devant ce problème ne laissant guère pressentir de solution, quelques études de laboratoires peuvent maintenant montrer la voie de nouvelles recherches.
Un aspect puisé dans des études plus anciennes de laboratoire est l’observation des effets ON et OFF (champ enclenché et champ supprimé). Comme effets de champs présents, les réponses cellulaires aux expositions aux champs électromagnétiques peuvent être éphémères, même si l’exposition est maintenue. Ceci fut observé pour la réponse de la protéine-kinase dans les lymphocytes humains aux champs de fréquences radio, modulées en E.L.F. Une diminution rapide de l’activité de 60 % en 15 à 30 minutes fut suivie d’un retour au niveau de celui des cellules-témoins en une heure, alors que l’application des champs continuait (Byus et al., 1984).
Dans des cellules osseuses exposées à des champs magnétiques pulsés 72 Hz utilisés dans les traitements de fractures, Luben et al. (1994) ont noté que les cellules osseuses sont désensibilisées aux effets de l’hormone parathyroïde (PTH) durant l’exposition, mais retrouvent leur sensibilité au moment de l’arrêt (OFF) d’application du champ. Luben et al. (1994) ont estimé que l’interférence avec la transduction du signal par l’intermédiaire de la membrane est
un mécanisme plausible, grâce auquel des champs magnétiques à faible énergie peuvent influencer des processus à l’intérieur des cellules. Leur étude a comparé l’action de champs magnétiques sinusoïdaux 60 Hz , 0,1 mT avec l’action du promoteur tumoral TPA. Les deux provoquent une augmentation éphémère de l’enzyme-messagère protéine-kinase C (PKC), mais réduisent l’activité dans le protoplasme cellulaire.
En arrêtant le champ (OFF), ils ont obtenu le retour du PKC, comme cela a pu être montré par une augmentation de 4 à 5 fois dans l’activité totale du PKC cellulaire sur une période de 4 heures. Durant cette période d’exposition, il y avait un maximum de désensibilisation du récepteur de l’hormone parathyroïde.
Dans un cadre de temps différent, les études épidémiologiques ont soulevé des questions à propos des effets potentiels sur la santé d’importantes variations de champs magnétiques et de lourdes charges de départ associées avec des opérations de mise en route d’appareils industriels et domestiques. Des études de laboratoire ont tenu compte du temps de cohérence, durant lequel un champ électromagnétique doit être maintenu à une fréquence E.L.F. spécifique, avant de basculer vers une autre fréquence différente mais relativement proche, en vue d’initier une réponse enzymatique. Litovitz et al. (1993) ont testé les réponses de l’ornithine décarboxylase (ODC) à des composantes électromagnétiques à extrêmement basses fréquences (E.L.F.) d’un champ électromagnétique enclenchant sur 55 ou sur 65 Hz. Ils ont montré que le temps de cohérence minimal pour cette réponse particulière est situé entre 1 et 10 secondes pour obtenir une réponse enzymatique fiable.
Il est clair que ces études ne sont que des recherches pilotes en vue de déterminer des mesures de valeurs d’exposition se rapportant à des effets sur la santé humaine. Cependant, elles mettent en exergue la grande complexité des effets des champs électromagnétiques non ionisants sur les tissus vivants. Contrairement aux radiations ionisantes, où les doses acceptables par les tissus peuvent être calculées de façon relativement simple, à partir des produits fournis par l’intensité de la radiation sur le tissu et de la durée de cette exposition, la métrologie des radiations non ionisantes doit tenir compte des effets profonds sur les tissus, attribuables à l’intermittence des expositions et au caractère fréquemment passager de la réponse biologique consécutive.
12. résumé et conclusions.
Au cours de ces quinze dernières années, des questions préoccupantes se sont posées quant à l’énergie électrique. L’usage important est sans cesse croissant de l’énergie électrique à travers le monde, avec ses incalculables bienfaits pour toute l’humanité, peut amener une quantité d’effets néfastes pour la santé. Au cours de cette même période, des recherches menées en vue d’évaluer les niveaux des dangers possibles ont suivi deux chemins, avec les études épidémiologiques et les études de laboratoires, progressant parallèlement, mais apportant peu d’options en vue d’une coordination.
L’étendue et le contenu de ces recherches de laboratoire en bioélectromagnétisme ont été significativement gonflées en étendue et en contenu, parce que leurs principaux financements encore que très réduits étaient apportés par les agences gouvernementales et consistaient en des mandats dans la recherche des risques.
En dépit de ces embarras, les recherches en bioélectromagnétisme semblent avoir dépassé les buts immédiats dans la recherche des risques, pour approcher d’une première compréhension la nature essentielle de la matière vivante en termes de processus physiques au niveau atomique, loin au delà du règne des réactions chimiques dans l’élaboration des biomolécules.
Les études de laboratoire ont identifié les membranes cellulaires comme sites tissulaires primaires d’interaction avec les champs électromagnétiques environnementaux. Elles ont déterminé les séquences principales dans le couplage des signaux à la surface des cellules avec une cascade de mécanismes enzymatiques à énergie élevée à l’intérieur des cellules, incluant les mécanismes régulant la croissance cellulaire. Ces études indiquent une conjonction entre les actions des promoteurs chimiques de cancers et les champs électromagnétiques au niveau de la membrane cellulaire comme une étape-clé dans la formation des tumeurs. Le rôle des radicaux libres dans la première détection des champs électromagnétiques à des niveaux athermiques est conforté par les modèles biophysiques et les données expérimentales.
D’une part, l’importance de ces nouvelles connaissances souligne l’émergence du bioélectromagnétisme comme un champ de recherches interdisciplinaires aux frontières de la physique et des sciences de la vie. Ces nouvelles voies portent un grand espoir pour des progrès futurs, dans la compréhension des fonctions du corps humain en bonne santé et malade. D’autre part, sans beaucoup d’autres recherches fondamentales, il n’existe qu’un faible espoir de développer des approches de valeurs mesurables pour les doses adaptées aux tissus vivants, en matière d’exposition aux champs électromagnétiques. Sans cette échelle de mesures, d’autres études épidémiologiques ne semblent devoir apporter qu’un faible espoir de progrès importants.
La dernière décade a montré une érosion progressive de l’évaluation normale des nouvelles connaissances par des experts qualifiés dans les domaines particuliers des sciences médicales et de la pratique médicale, via des revues examinées par des pairs et via des échanges collégiaux. A la place, il y a le spectre troublant de la corporation des hommes de lois importuns par leurs poursuites de la recherche toujours en cours, en épousant une cause litigieuse plutôt qu’en cherchant à découvrir un système légal destiné à établir une crédibilité scientifique. Ceci peut s’appuyer sur l’exemple de la publicité accordée dans les salles d’audience aux distorsions de connaissances méticuleusement acquises en matière d’interactions entre le corps humain et les champs électromagnétiques ambiants.
Il y a plus de 1900 ans, Tacite connaissait bien le destin de la société romaine qui s’était abandonnée aux machinations des hommes de loi : Si personne ne paie les procès, il y en aura moins! Ainsi, les querelles, les recherches de chefs d’accusations, les malveillances et les diffamations sont encouragés. Car tout comme une maladie physique est une source de revenus pour le médecin, un système légal malade enrichit les avocats.
************************
note du traducteur
W.R.Adey a publié ce texte en 1994. Il existe indiscutablement un fossé entre le début de l’article où les effets de champs sur les cellules et les animaux de laboratoire sont décrits et la fin de l’article ainsi que les conclusions.Or, en juin-juillet 1995 un article de Microwave News faisait référence à une indiscrétion à propos de propositions formulées par le Conseil National Américain de Protection (N.C.R.P.) contre les Radiations en vue de normaliser dans le futur, les expositions aux champs électromagnétiques 60 Hz. Ce conseil, présidé par W.R. Adey propose de limiter à 2 mG et à 10 V/m l’exposition générale du public dans les 10 années à venir. Il assortit cette recommandation d’une tendance progressive vers une norme A.L.A.R.A. (Aussi faible qu’il est raisonnablement possible de le réaliser), comme pour les radiations nucléaires. Ce conseil doit présenter cette proposition à l’Environmental Protection Agency, dont il est l’émanation. Cette norme est 500 fois inférieure aux recommandations de l'I.R.P.A. Le revirement de W.R. Adey, suite aux nombreuses études publiées est évident.
La critique formulée dans cet article à l’intention des hommes de loi nous parait déplacée; en effet il faut comprendre le sort des populations contraintes arbitrairement de vivre sous des lignes à haute tension ou à proximité d’antennes émettrices. La civilisation industrielle ne fait que très peu de cas des problèmes de santé que ces citoyens rencontrent et les habitants de certaines régions sont livrés à un arbitraire qui ressemble fort à une attitude dictatoriale. Ils sont pour ainsi dire sacrifiés à un supposé intérêt public. Ils finissent par se considérer comme des citoyens de seconde catégorie, comme des rebuts. Si les états, face aux lobbies industriels refusent de tenir compte des avis de la population, où est la démocratie, où est le droit constitutionnel ? Dans ces situations qui révèlent effectivement la maladie des institution , il est normal que les hommes de loi s’érigent en protecteurs des citoyens contre l’état! Il s’agit en fait d’une lutte contre une forme plus sournoise de corruption.
références bibliographiques
- adey w. r., Tissue interactions with nonionizing electromagnetic fields, in Physiol. Rev, n. 61, pp. 435-514 (1981).
- adey w. r., Joint actions of environmental nonionizing electromagnetic fields and chemical pollution in cancer promotion, in Environmental Health Perspectives, n. 86, pp. 297-305 (1990).
- adey w. r., Electromagnetics in biology and medicine, in Modern Radio Science, h. matsumoto, Oxford University Press, pp. 227-245 (1993).
- adey w. r., Extremely low frequency magnetic fields and promotion of cancer, in Interaction mechanisms of low level electromagnetic fields in living systems, b. norden, c. ramel, Oxford University Press, pp. 23-46 (1992).
- adey w. r., lawrence a.f., Nonlinear electrodynamics in biological systems, New York, Plenum Press (1984).
- bawin s.m., adey w. r., Sensitivity of calcium binding in cerebral tissue to weak electric fields oscillating at low frequency, in Proc. Nat. Acad. Sci. USA, n. 73, pp. 1999-2003 (1976).
- bawin s.m., kaczmarek l.k., adey w.r., Effects of modulated VHF fields on the central nervous system, Ann. NY Acad. Sci., n. 247, pp. 74-80 (1975).
- bawin s.m., satmary w.m., adey w.r., Nitric oxide modulates rythmic slow activity in rat hippocampal slices, in Neuro Report Volume, n. 5 (15) (1994).
- berridge m.j., Inositol triphosphate and diacylglycerol: two interacting second messengers, in Ann. Rev. Biochem., n. 56, pp. 159-193 (1987).
- blackman c.f., Effect of electrical and magnetic fields on the nervous system, in The Vulnerable Brain and Environmental Risks. Vol. 3. Toxins in air and water, r.l. isaacson et k. f. jensen, New York, Plenum Press, pp. 341à 355 (1994).
- bignami m., rosa s., falcone g., tato f., katoh f., yamasaki h., Specific viral oncogenes cause differential effects in cell-to-cell communication, relevant to suppression of the transformed phenotype by normal cells, in Molec. Carcin., n. 191, pp.67-75 (1988).
- blackman c.f., elder j.a., weil c.m., benane s.g., eichinger d.c., house d.e., Induction of calcium ion flux from brain tissue by radio frequency radiation - Effects of modulation frequency and field strength, in Radio Sci., n. 14, pp.93-98. (1979).
- blackman c.f., benane s.g., house d.e., joines w.t., Effects of ELF (1-120 Hz) and modulated (50 Hz) RF fields on the efflux of calcium ions from brain tissues, in vitro, in Bioelectromagnetics, n. 6, pp.1-12 (1985a).
- blackman c.f., benane s.g., rabinowitz j.r., house d.e., A role for the magnetic field in the radiation-induced efflux from brain tissue, in vitro, in Bioelectromagnetics, n. 6, pp.327-338 (1985b).
- byus c.v., lundak r.l., fletcher r.m., adey w.r., Alterations in proteine kinase activity following exposure of cultured lymphocytes to modulated microwave fields, in Bioelectromagnetics, n. 5, pp.34-51 (1984).
- byus c.v., pieper s.e., adey w.r., The effects of low energy 60 Hz environmental electromagnetic fields upon the growth-related enzyme ornithine decarboxylase, in Carcinogenesis, n. 8, pp.1385-1389 (1987).
- byus c.v., kartun k., pieper s., adey w.r., Increased ornithine decarboxylase activity in cultured cells exposed to low energy modulated microwave fields and phorbol ester tumor promoters, in Cancer Res., n. 48, pp. 4222-4226 (1988).
- byus c.v., Regulation of the efflux of putrescine and cadaverine from rapidly-growing cultured RAW-264 cells by extracellular putrescine, in Biochem. J., in press (1994).
- cain c.d., thomas d.l., adey w.r., 60 Hz magnetic field acts as a co-promoter in focus formation of C3H10T1/2 cells, in Carcinogenesis, n. 14, pp. 955-960 (1993).
- castagna m., takai y., kaibuchi k., sano k., kikkawa u., nishizuka y., Direct activation of calcium-activated phospholipid-dependant protein kinase by tumor promoting phorbol esters, in J. Biol. Chem., n. 257, pp.7847-7851 (1982).
- conti p., gigante g.e., alesse e., cifone m.g., fieschi c., angeletti p.u., Effect of electromagnetic field on two calcium dependant biological systems, in J. Bioelectr., n. 4, pp. 227-236 (1985b).
- dutta s.k., subramoniam a., ghosh b., parshad r., Microwave radiation-induced calcium efflux from brain tissue, in vitro, in Bioelectromagnetics, n. 5, pp.71-78 (1984).
- foster k.r., guy a.w., The microwave problem., in Sci. Amer., n. 225 (3), pp.32-39 (1986).
- foster k.r., pickard w.f., Microwave: the risks of risk research, in Nature London, n. 350, pp.531-532 (1987).
- frohlich h. ed., Biological coherence and response to external stimuli, Heidelberg, Springer (1988).
- grundler w., kaiser f., keilmann f., walleczek j., Mechanics of electromagnetic interaction with cellular systems, in Naturwissenschaften, n. 79, pp.551-559 (1992).
- harkins t.t., grissom c.b., Magnetic field effects on B12 ethanolamine ammonia lysase : evidence of a radical mechanism,in Science, n. 263, pp.958-960 (1994).
- hill s.m., blask d.e., Effects on the pineal hormone melatoni on the proliferation and morphological characteristics of human breast cancer cells (MCF-7) in culture, in Cancer Res., n. 48, pp. 6121-6126 (1988).
- kaiser f., Theory of non linear excitation, in Biological coherence and response to external stimuli, frohlich h., Heidelberg, Springer, pp. 25-48 (1988).
- liboff a.r., The cyclotron resonance hypothesis: experimental evidence and theorical constraint, in b. norden, c. ramel, Oxford University Press, p 130-147 (1992).
- lidbury r.p., harland j.d., hefferman c., seeley m., dunham e.e., Inhibition of melatonin?s natural oncostatic action on MCF-7 cells : 60 Hz dose threshold determination, in Bioelectromagnetic Society. 16th Annual Meeting, Copenhagen, Proceedings, pp.51 (abstract) (1994).
- lin-liu s., adey w.r., Low frequency amplitude modulated microwave fields change calcium efflux rates from synaptosomes, in Bioelectromagnetics, n. 3, pp. 309-322 (1982).
- litovitz t.a., krause d., penafiel m., elson e.c., mullins j.m., The role of coherence time in the effect of microwaves on ODC activity., in Bioelectromagnetics, n. 14, pp. 395-403 (1993).
- loewenstein w.r., Junctional intercellular communication: the cell-to-cell communication channel, in Physiol. Rev., n. 6, pp.829-913 (1981).
- luben r.a., Effects of low energy electromagnetic fields (pulsed and DC) on membrane signal transduction processes in biological systems, in Health Physics, n. 61, pp. 15-28 (1989).
- luben r.a., cain c.d., Use of hormone receptor activities to investigate the membrane effects of low energy electromagnetic fields, in Nonlinear Electrodynamics in Biological Systems, w.r. adey et a.f. lawrence, New York, Plenum Press, pp. 23-34 (1984).
- luben r.a., cain c.d., chen m-y., rosen d.m., adey w.r., Effects of electromagnetic stimuli on bone and bone cells, in vitro: inhibition of response to parathyroid hormone by low-frequency energy fields, in Proc. Natl. Acad. Sc. USA, n. 79, pp. 4180-4183 (1982).
- luben r.a., hyunh d., weinshank r.l., smith l.e., Molecular cloning of candidate sequences for the mouse osteoblast parathyroidhormone receptor, in Calcium Regulation and Bone Metabolism, d.v. cohn, f.h. glorieux, t.j. martin , Amsterdam, Elsevier, pp. 39-44 (1990).
- luben r.a., morgan a.p., carlson a., duong m., One gauss 60 Hz magnetic fields modulate protein kinase activity by a mechanism similar to that of tumor promoting phorbol esters, in Bioelectromagnetics Society. 16th. Annual Meeting, Copenhagen, proceedings, pp.74 (1994).
- lyle d.b., ayotte r.d., sheppard a.r., adey w.r., Suppression of T lymphocyte cytotoxicity following exposure to 60 Hz sinusoidal electric fields, in Bioelectromagnetics, n. 9, pp. 303-313 (1988).
- lyle d.b., schechter p., adey w.r., lundak r.l., Suppression of T lymphocyte cytotoxicity following exposure to sinusoidally amplitude modulated fields, in Bioelectromagnetics, n. 4, pp.281-292 (1983).
- lyle d.b., wang x., ayotte r.d., sheppard a.r., adey w.r., Calcium uptake by leukemic and normal T lymphocytes exposed to low frequency magnetic fields, in Bioelectromagnetics, n. 12, pp. 145-156 (1991).
- mc bain c.j, mayer m.l., N-methyl-D-aspartic acid receptor structure and function, in Physiol. Rev., n. 74, pp. 723-760 (1994).
- mc lauchlan k., Are environmental electromagnetic fields dangerous ?, in Physics World, pp. 41-45 (Janvier 1992).
- mc lean j.r.n., stuchly m.a., mitchel r.e.j., wilkinson d., yang h., goddard m., lecuyer d.w., schunk m., callary e., morrisson d., Cancer promotion in a mouse-skin model by a 60 Hz magnetic field. Tumor development and immune response, in Bioelectromagnetics, n. 12, pp. 273-288 (1991).
- miura m., takayama k., okada j., Increase in nitric oxide and cyclic-GMP of rat cerebellum by radiofrequency burst-type electromagnetic radiation, in J.Physiol. London , n. 461, pp.513-524 (1993).
- nishizuka y, Protein kinase C as a possible receptor protein of tumor-promoting phorbol ester, in J. Biol. Chem., n. 258, pp. 11442-11446 (1983).
- nishizuka y., The role of protein kinase C in cell surface signal transduction and tumour promotion, in Nature London, n. 308, pp. 693-698 (1984).
- phillips j.l., Effects of EM field exposure on gene transcription, in J. Cell. Biochem., n. 51, pp.381-386. (1993).
- phillips j.l., haggren w., thomas w.t., ishida-jones t., adey w.r., Magnetic field-induced changes in specific gene transcription, in Biochim. Biophys. Acta, n. 1132, pp.140-144 (1992).
- pitot h.c., dragan y.p., Facts and theories concerning the mechanisms of carcinogenesis, in FASEB J., n. 5, pp. 2280-2288 (1991).
- reiter r.j., Alterations of the circadian melatonin rhythm by the electromagnetic spectrum : a study in environmental toxicology, in Regulatory Toxicology and Pharmacology, n. 15, pp. 226-244 (1992).
- schumann w.o., Uber elektrische Eigenschwingungen des Hohlraumes Erd-Luft-Ionosphäre, erregt durch Blitzentladungen, in Zeitschrift. Angew. J. Phys., n. 9, pp. 373-378 (1992).
- semm p.,Neurobiological investigations on the magnetic sensitivity of the pineal gland in rodents and pigeons, in Comp. Biol. Physiol., n. 159, pp. 619-625 (1983).
- slaga t.j., sivak a., bouwell r.k., Mechanisms of tumor promotion and carcinogenesis, vol.2, New York, Raven Press (1978).
- steiner u.e., ulrich th., Magnetic field effects in chemical kinetics and related phenomena, in Chem. Rev., n. 89, pp.51-147 (1989).
- tabib a., bachrach u., Activation of the proto-oncogene c-myc and c-fos by c-ras: involvement of polyamines, in Biochem. Biophys. Res. Communications, n. 202, pp.720-727 (1994).
- terada h., kitagawa f., okamoto n., watanabe s., taki m., saito m., An analysis of dose in tissue irradiated by near field of a circular loop antenna, in IEICE Trans. Commun., n. E77-B, pp. 754-761 (1994).
- walleczek j., Electromagnetic field effects on cells of the immune system: the role of calcium signaling, in FASEB J., n. 6, pp. 3176-3185 (1992).
- walleczek j., Immune cell interactions with extremely low frequency magnetic fields: experimental verification and free radical mechanisms, in On the nature of electromagnetic fields interactions with biological systems, a.h. frey, New York, R.G. Landes Company (1994).
- walleczek j., lidbury r.p., Nonthermal 60 Hz sinusoidal magnetic field exposure enhances 45Ca2+ uptake in rat thymocytes: dependance on mitogen activation, in FEBS Lett., n. 271, pp. 157-160 (1990).
- walleczek j., killoran p.l., adey w.r., Acute 60 Hz magnetic field effect on Ca2+ influx in human Jurkat T-cells: strict dependence on cell state, in Bioelectromagnetics Society. 16th. Annual Meeting, Copenhagen. Proceedings, p.76 (abstract) (1994).
- weinstein i.b., The origins of human cancer : molecular mechanisms of carcinogenesis and their implications for cancer prevention and treatment, in Cancer Res., n. 48, pp.4135-4143 (1988).
- wilson b.w., anderson l.e., hilton d.i., phillips r.d., Chronic exposure to 60 Hz electric fields: effects on pineal functions in the rat, in Bioelectromagnetics, n. 2, pp. 371-380 (1981).
- wilson b.w., anderson l.e., ELF electromagnetic-field effects on the pineal gland, in Extremely low frequency electromagnetic fields : The question of cancer, b.w. wilson, r.g. stevens, l.e. anderson, Columbus, Ohio, Battelle Press, pp. 159-186 (1990).
- yamasaki h., The role of cell-to-cell communication in tumor promotion, in Nongenotoxic mechanisms in carcinogenesis, t.e. butterworth, t.j. slaga, 25th. Banbury Report, Cold Spring Harbor Laboratory (1987).
- yamasaki h., Aberrant expression and function of gap junctions during carcinogenesis, in Environ. Health Perspectives, n. 93, pp. 191-197 (1991).
- yellon s.m., Acute 60 Hz magnetic field exposure effects on the melatonin rhythm in the pineal gland and circulation of the adult Djungarian hamster, in J. Pineal Res., n. 16, pp.136-144 (1994).