Les champs �lectriques, vecteurs d'informations biologiques
Note introductive de J.M. Danze
����������� Il
y a quelques ann�es, � une �poque o� la t�l�phonie mobile n'avait pas encore
pris possession de notre vie, parfois contre le gr� de certains de nos concitoyens,
le Prof. Urlrich Warnke, de l'Universit� de Saarbrucken m'a remis une premi�re
mouture en langue anglaise de ce texte. Je n'y avais alors gu�re attach� d'importance,
si ce n'est dans un cadre scientifique pur. Puis, dans la suite, Ulrich Warnke
est devenu l'un de mes ma�tres � penser et j'ai d�couvert une deuxi�me mouture�
plus �volu�e de ce texte. Je l'avais alors transmise telle quelle � un cercle
apicole belge en vue d'une traduction r�sum�e pour publication. Il y a beaucoup
de chance que l'importance de ce texte soit pass�e au del� de l'esprit des
dirigeants de ce groupe, car aucun �cho n'y a �t� fait.
����������� Aujourd'hui, � l'heure o� les t�l�phones mobiles
et leurs antennes relais parasitent certains appareils m�dicaux et le
fonctionnement des radio-t�lescopes, il serait peut-�tre bon de se demander
s'ils ne sont pas en train �galement de toucher le monde des insectes
butineurs (abeilles, bourdons ) et des oiseaux (moineaux, �tourneaux etc�).
����������� J'ai donc d�cid� de traduire ce dernier texte dans
son int�gralit� afin de permettre (je l'esp�re) une meilleure compr�hension
de ces ph�nom�nes d'informations biologiques par champs �lectriques interpos�s.
����������� J'esp�re que les entomologistes et les ornithologues
daigneront prendre connaissance de ce document pour confronter les donn�es
de U. Warnke avec des observations cons�cutives dans le domaine des champs
alternatifs 50 Hz et surtout des micro-ondes de t�l�phonie mobile�
����������� Il est peut �tre grand temps !
Les champs �lectriques, vecteurs d'informations biologiques�
������ par
le Prof. Ulrich WARNKE
������ Institut de Biologie, Universit� de Saarbr�cken
Convergence
des champs biologiques dans l'eau et sur terre
Charge
�lectrostatique des animaux
Conductivit�
de la surface du corps
Charge de base
par �lectricit� de contact
Charge et d�charge
gr�ce aux composantes �lectriques atmosph�riques
Animaux
en vol
Animaux
au sol
Pi�zo-�lectricit�,
pyro-�lectricit� et ph�nom�nes d'�lectret
Ph�nom�nes
photo-�lectriques
Implication
biologique des champs �lectriques
Le
facteur de stress
Apprentissage
Transporteurs
d'informations
Les
r�f�rences de temps
Electrocommunication
Vol�es d'oiseaux
Relations
entre les forces �lectrostatiques en vol en formation
G�om�trie
calcul�e de la formation
V�rification
de l'hypoth�se sur base de documents photographiques de formations de
vol
Electro-orientation
et �lectronavigation
M�canismes
de perception des champs �lectriques � extr�mement basses fr�quences (de
1 Hz � 3 kHz) et � tr�s basses fr�quences (de 3 kHz � 30 kHz).
Microvibrations
Effets
de pointes
Convergence des champs biologiques dans l'eau et sur terre
����������� Des
poissons d�veloppant de puissantes et de faibles d�charges �lectriques sont
maintenant bien connus des experts. Cependant, le fait que tous les poissons,
y compris ceux que l'on consid�re comme non �lectriques, sont entour�s d'un
champ continu dipolaire (� � la bouche; + aux branchies) modul� par la respiration
m�caniques est moins connu. Ce champ a une signification biologique et est
utilis� par des esp�ces � faible �volution phylog�nique, comme les requins,
les raies et les esturgeons, comme un signe destin� � la pr�dation. Une plie
dissimul�e sur le fond de la mer, par exemple, peut ainsi �tre localis�e par
des pr�dateurs passant � proximit�, gr�ce � son champ �lectrique. Les ordres
de poissons d'eau douce et d'eau sal�e � �volution plus �lev�e ont apparemment
d�velopp� leur utilisation des champs �lectriques pour y introduire une communication
(perception) inter-esp�ces ainsi que la localisation de mati�res (activation).
Les bancs migrateurs de poissons semblent �tre maintenus ensemble par leur
champ �lectrique commun et ils utilisent probablement certains niveaux de
potentiel dans la mer, en vue de leur navigation. La quantit� isol�e d'�nergie
n�cessaire pour initier une action de perception est �tonnamment basse.
����������� On a jusqu'ici totalement ignor� que des champs
�lectriques sont �galement g�n�r�s par des animaux terrestres; de nombreuses
esp�ces d'insectes, d'oiseaux et d'animaux � fourrure atteignent des amplitudes
de champ relativement �lev�es [37,
38]. Sur terre ces champs biologiques sont un facteur impliqu�
dans la physiologie.
����������� Ceci nous am�ne � la question int�ressante de savoir
� quel point des ph�nom�nes �lectriques convergents, d�j� �tablis comme des
moyens effectifs d'�change de communications dans l'eau se sont �galement
d�velopp�s sur terre.
����������� Il faut se souvenir de ce qu'au moment de l'�mergence
des premiers organismes terrestres, les champs �lectriques et magn�tiques
atmosph�riques, les oscillations �lectromagn�tiques et les ions atmosph�riques
�taient d�j� des facteurs m�t�orologiques ou extra-terrestres corr�l�s et
quantifi�s parmi l'espace de vie. Il semble plut�t invraisemblable que ces
conditions fondamentales ne fussent pas exploit�es par les organismes dans
un but informationnel. L'importance essentielle du codage �lectrique en vue
de la m�morisation des informations endog�nes et de l'�change d'informations
est bien connue et de l� l'utilisation d'�nergie �lectrique ou magn�tique
endog�ne devrait �galement �tre consid�r�e comme une source r�gulatrice d'informations.
Ceci pourrait jouer in r�le particuli�rement important chaque fois que des
stimuli via des canaux de sens plus hautement d�velopp�s ne re�oivent plus
d'information, comme c'est le cas par exemple, d'oiseaux migrateurs volant
� haute altitude la nuit ou au dessus de grandes �tendues oc�aniques.
����������� Ce sont autant d'�l�ments pour l'hypoth�se
de travail.
Charge �lectrostatique des animaux
����������� Lorsqu'une
plume est agit�e dans l'air, elle se charge d'�lectricit� statique, pourvu
que des porteurs de charge soient pr�sents dans l'air.
����������� Lorsqu'un oiseau vole dans l'air, cependant ou m�me s'il est pos� sur
une branche, les processus deviennent un peu plus complexes. Dans des conditions
"naturelles" il y a beaucoup de variables interd�pendantes en interrelations
impliqu�es et celles-ci doivent �tre soumises � des �valuations quantitatives
et �tre enregistr�es individuellement quant � leur magnitude et � leur d�pendance
vis-�-vis du facteur temps.
����������� Cependant, cette t�che d�passe de loin les possibilit�s des tests disponibles
et par cons�quent, les animaux et les mod�les sont explor�s dans un laboratoire
en fonction de param�tres d�finis de variables qui simulent les conditions
physiologiques. Les r�sultats d�montrent les principes et les facteurs d�pendants
de la charge �lectrostatique des insectes, des oiseaux et des mammif�res,
mais ne peuvent fournir que des valeurs approchant les valeurs absolues.
����������� L'ordre selon lequel les ph�nom�nes r�pondant aux g�n�rateurs de champs
biologiques sur terre sont d�crits correspond � leur fr�quence d'apparition.
Conductivit� de la surface du corps
����������� Les champs �lectriques � amplitudes plus �lev�es ne peuvent �tre mis en �vidence chez des animaux que si les charges unipolaires divis�es (la cause de ces champs �lectriques) ne sont pas continuellement neutralis�es. La neutralisation des charges est principalement une fonction de la mobilit� des charges � la fois sur la surface du corps et � l'int�rieur du corps.
����������� Tous
les insectes terrestres ayant une cuticule et toutes les esp�ces animales
� �piderme squameux, � coquille, � plumes ou � fourrure sont prot�g�s gr�ce
� ces organes cutan�s poss�dant des surfaces isolantes excellentes (R�sistivit�
sp�cifique de 1010 W/cm � 1012 W/cm).
����������� La m�thode suivante de d�termination des
variations de la r�sistance relative locale sur la surface du corps: "l'int�rieur"
d'un animal vivant fixe acquiert un potentiel via la bouche; une abeille,
par exemple, qui re�oit un m�lange de miel et d'eau pr�alablement charg�e
� un potentiel oppos� � celui de la terre. Deux mesures s�par�es de r�sistances
d�terminent alors l'imp�dance de la source prot�g�e par la couche externe
du corps contre la d�charge de l'int�rieur du corps. Des valeurs absolues
ne peuvent �tre obtenues qu'in vitro au moyen d'un pont de mesures (pont
de Wheatstone), � partir d'�chantillons de tissus sp�cifiques. Un microscope
�lectronique � balayage peut fournir une pr�sentation optique r�elle des
zones de conductivit�s diff�rentes sur des plaquettes de pr�parations
virtuellement fra�ches et non trait�es, sur base d'un feuillet d'�chantillon
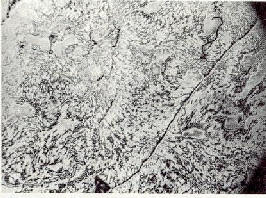
(fig.1).
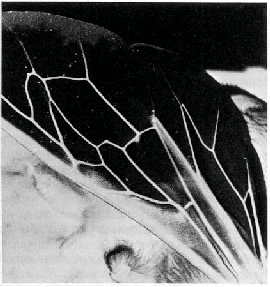 Fig.1: Image d'une aile d'insecte en test. La modification de la mosa�que de conductivit� peut �tre habituellement suivie sous un microscope �lectronique et d�pend de la longueur de la p�riode sous vide. Des structures de lumi�re l�g�rement color�es indiquent des densit�s de courant relativement �lev�es, c'est-�-dire une conductivit� �lev�e. Les zones sombres indiquent le contraire. Etant donn� que le fluide capillaire s'ass�che, les veinules - ici toujours lumineuses -� deviennent progressivement sombres. |
La
surface du corps de toute esp�ce animale pr�sente une mosa�que typique
de zones de conductivit�s diff�rentes. Cette mosa�que est sp�cifique et
clairement dessin�e; elle concorde virtuellement avec le profil des charges
en superficie. Des modifications dans cette disposition sont d�termin�es
soit par des s�cr�tions �lectriquement actives, soit par des param�tres
physiques environnementaux. Une humidit� relative �lev�e aboutit ainsi
� des valeurs de conductivit� �lev�e sur des zones isol�es de la surface
du corps lesquelles r�sultent du d�p�t d'un film adh�rent d'humidit�.
La chaleur ajout�e provoque une augmentation nette de la conductivit�
sur toutes les surfaces du corps mentionn�es ci-dessus, en raison des
principes r�gissant les �lectrolytes et les semi-conducteurs. Des modifications
de l'intensit� lumineuse et des diff�rences de concentrations en ions
atmosph�riques exercent �galement une influence, bien qu'� un niveau limit�
(Fig. 2).
����������� Il n'y a pas de doute que les surfaces
organiques du corps aient des propri�t�s semi-conductrices, mais la preuve
exp�rimentale est difficile � obtenir. La conductivit� ionique pr�domine
dans les tissus frais; elle est due au pourcentage �lev� d'eau pr�sente
et dissimule donc les effets semi-conducteurs.
����������� D'autre part, la d�shydratation d�truit l'�tat
fonctionnel biologique normal du tissus. L'effet redresseur de courant
commun�ment trouv� dans les semi-conducteurs ne peut �tre utilis� ici
comme indicateur � cause du contact in�vitable entre le mat�riau de l'�lectrode
et la substance organique, lequel contact fausse les transitions n-p.
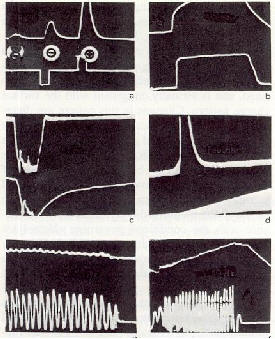 Fig.2: Oscillogrammes de l'augmentation induite de la conductivit� sur la cuticule de l'insecte (trac� sup�rieur). Des r�sultats analogues apparaissent dans des �chantillons de k�ratine, de collag�ne et de dentine. Le trac� inf�rieur montre le signal artificiel. a): concentration ionique atmosph�rique bipolaire et unipolaire augment�e. b): temp�rature augment�e. c): temp�rature diminu�e. d): humidit� absolue augment�e. e) et f): variations de l'intensit� de la lumi�re [1]. |
����������� Si
l'on consid�re la charge statique des animaux terrestres, il est important
de tenir compte non seulement de la conductivit� normale de la surface du
corps, mais surtout de la nature des surfaces via lesquelles a lieu le contact
avec la terre. Les animaux qui poss�dent des glandes sudoripares ou des glandes
odorif�res ou des glandes adh�sives aux endroits par lesquels ils sont en
contact avec la surface de parcours sont donc �quip�s d'un excellent contact
galvanique, alors que ceux qui marchent sur des sabots, sur des doigts ou
sur des griffes sont presque enti�rement isol�s de la terre. Un fait int�ressant
et inhabituel se manifeste chez beaucoup d'esp�ces d'insectes : les mouches,
les abeilles et d'autres insectes poss�dent un rabat adh�sif (arolium)
entre deux griffes de leurs pattes. Ce rabat adh�sif peut �tre d�ploy� ou
contract� � tout moment lorsque l'insecte est au sol. Lorsque l'arolium est
en contraction, l'insecte rampant sur ses griffes est ainsi isol� �lectriquement
de l'environnement et est capable de recevoir une charge statique. Si d'autre
part le rabat adh�sif d�ploy� touche la surface de parcours, l'insecte se
met rapidement au potentiel de cette surface. Pour certaines parties de l'insecte,
ceci peut impliquer une d�charge ou une modification de la charge, parfois
avec un changement de polarit�.
����������� Les animaux a�riens sont fortement isol�s �lectriquement,
en fonction d'une r�sistivit� atmosph�rique moyenne Ra = 1015
W/cm.
����������� En ce qui concerne la conductivit� superficielle, l'�tre humain occupe
une position particuli�re � cause de la r�partition � forte densit� des glandes
sudoripares sur l'ensemble du corps. Des modifications de temp�rature, des
variations d'humidit� relative et des facteurs psychologiques peuvent �tre
la cause de consid�rables variations de la r�sistance ohmique. Les fluctuations
de l'activit� glandulaire sont impliqu�es dans ces ph�nom�nes. C'est pourquoi
une charge statique plus �lev�e accompagn�e d'un contact galvanique avec la
terre n'est possible que par les poils, les cheveux en particulier. D'autre
part, la polarisation di�lectrique est d�tectable sur toutes les parties du
corps.
Charge
de base par �lectricit� de contact
�����������
Les insectes et les oiseaux re�oivent leur
charge de base par �lectricit� de contact: deux processus doivent �tre
consid�r�s s�par�ment:
- la tension (voltage) de contact entre le mat�riau de la surface de l'animal (k�ratine, chitine, substances cireuses) et les couches d'air environnantes,
- la tension (voltage) de contact entre plusieurs structures du corps
�����������
Pendant le contact de deux surfaces, jusqu'� une distance mol�culaire
(10-10 m), la s�paration des charges positives et n�gatives a lieu
par le transfert d'une certaine quantit� d'�nergie aux points de contact.
L'importance de la modification des charges d�pend de trois variables physiques
du mat�riau selon les r�gles d�finies: l'�nergie d'ionisation, le travail
de lib�ration thermique des �lectrons et l'affinit� des �lectrons. En cas
de s�paration des surfaces, l'une par rapport � l'autre, la surcharge locale
est maintenue pendant un temps plus long si la r�sistance de la surface est
inf�rieure � 1010 W. Lors
de la friction, de plus grandes parties de la surface sont amen�es en contact
�troit malgr� leur in�vitable rugosit� m�canique et des charges plus �lev�es
s'�laborent. L'�lectricit� obtenue par friction est l'une des plus anciennes
observations de l'humanit� et a donn� son nom � l'ensemble du ph�nom�ne (en
grec, Elektron signifie l'ambre). C'est pourquoi il est surprenant que l'�lectricit�
animale dans le milieu naturel ait � peine retenu l'attention jusqu'ici.
����������� En consid�rant maintenant l'�lectricit� de contact entre structures
d'un corps isol�, nous obtenons la description g�n�rale suivante [17,
24, 37].
La structure de la surface du corps � constante di�lectrique plus �lev�e
se charge positivement � l'oppos� de celle poss�dant une constante di�lectrique
plus faible. Les surfaces couvertes de poils se chargent positivement
en contact avec des surfaces glabres ou �cailleuses, lesquelles se chargent
alors n�gativement. Des structures corporelles similaires ne se chargent
que si les plans de contact ont des dimensions diff�rentes. Dans ce processus,
le plan le plus �tendu se charge positivement.
����������� Une quantit� de ph�nom�nes mesurables devraient �tre d�montr�s comme
des exemples dans ce cadre. En vol, les plumes d'habillage des oiseaux re�oivent
une charge positive par friction contre les plumes du duvet, ces derni�res
acqu�rant par contact une charge n�gative (Q = 10-9 As). Les oiseaux
ont l'habitude de lisser leurs plumes d'ailes en les faisant glisser dans
leur bec. Les plumes d'ailes sont ainsi charg�es positivement (Q = 10-10
As). Les abeilles lissent r�guli�rement leurs antennes avec un cr�neau de
nettoyage (nettoyeur d'antenne) aux pattes ant�rieures. L'antenne est ainsi
charg�e positivement (P = 1,3 Volt). Si les souris nettoient des parties de
leur corps avec leurs deux pattes de devant, elles re�oivent une surcharge
positive (P = 1 Volt).
����������� La tension de contact n'a une valeur reproductible que si le processus
� lieu sous des conditions constantes. Si les param�tres changent, et sp�cialement
la teneur en vapeur d'eau de l'air, les tensions prennent des valeurs tr�s
diff�rentes et peuvent m�me changer de signe 1.
����������� En ce qui concerne les corps d'animaux
isol�s �lectriquement, la tension de contact atteint souvent des valeurs �lev�es.
Tous les porteurs de charges dans l'air environnant, transport�s sur le site,
peuvent �tre attir�s ou repouss�s dans le m�me laps de temps gr�ce aux forces
de Coulomb rayonn�es par l'animal sous les conditions d�crites. Dans ce cas,
le courant de d�charge ne peut plus se produire et est � un niveau de saturation.
Si la production d'�lectricit� n'est pas interrompue � ce stade, il y aura
des surcharges �lev�es qui pourront provoquer des stimulations m�caniques-physiologiques
� cause du d�veloppement corr�l� et tr�s puissant de forces sur les plus petites
zones, par exemple entre les scl�rites et les membranes de la cuticule d'insectes
ou entre l'�lectrolyte du nez et l'�piderme mammaire.
����������� La composante d'�lectricit� de contact qui appara�t � partir du contact
entre l'animal et l'air est particuli�rement importante. Deux �tats cruciaux
doivent �tre distingu�s: l'animal au repos et l'animal en vol. Pour l'animal
au repos, la couche limite de la tension de contact se trouve dans l'air atmosph�rique
jusqu'� plusieurs rayons; il y a une tension spatiale diffus�e s'�laborant
autour de l'animal qui stimule la neutralit� �lectrique � l'ext�rieur.
����������� Cependant, cette situation change profond�ment si
des forces externes influencent ce syst�me d'�quilibre. Dans le cas de l'animal
volant, beaucoup de forces perturbatrices interf�rent fortement en m�me temps:
- un courant dans la phase d'air relatif au plan limite
- un champ �lectrostatique compos� de variables �lectriques a�riennes exog�nes et l'�lectricit� accroch�e au corps comme le g�n�rateur � plumes d�crit,
- des turbulences avec perforation compl�te des couches �lectriques en divers endroits in�gaux des surfaces (p. exemple des g�n�rateurs � vortex, fig. 23).
����������� Dans
tous ces cas, les porteurs de charges et particuli�rement ceux des parties
diffuses de la double couche sont d�cal�s et le plan limite pr�sente continuellement
des charges non satur�es. Un potentiel �lectrocin�tique calculable avec un
effet �lev� de g�n�rateur appara�t. Un �quilibre est atteint si le courant
�lectrique de retour compense exactement le courant de conversion entre les
nuages de charges isol�es dans l'espace. La cons�quence des �tats �lev�s de
r�sistance en ce qui concerne l'air d'une part et la surface de l'insecte
ou des plumes de l'oiseau d'autre part aboutit � ce que les processus de relaxation
aient une inertie extr�me et de ce fait, le niveau de champs de forces devient
tr�s �lev� (l'exemple d'un pigeon mesur� dans un tunnel a�rodynamique avec
de l'air ionis� a indiqu� E > 1000 V/m).
����������� Etant donn� que les forces perturbatrices
des doubles couches �lectriques d�crites atteignent leur expression maximale
aux ailes des animaux, les ailes des insectes volants et des oiseaux coupl�es
avec leur environnement d'air fonctionnent comme des g�n�rateurs de champs
�lectriques qui cr�ent de relativement puissantes forces de Coulomb � l'ext�rieur.
Charge
et d�charge gr�ce aux composantes �lectriques atmosph�riques
�����������
Fondamentalement, il y a deux composantes
de l'�lectricit� atmosph�rique qui son d'importance capitale dans la charge
et le d�charge des �tres humains, des animaux et des plantes. Elles sont:
- le gradient de potentiel
- la densit� de charge spatiale qui r�sulte d'une diff�rence entre les charges positives et n�gatives dans une unit� de volume donn�e.��������
Etant
donn� que ces valeurs servent probablement de signaux pour les animaux, il
en d�coule un bref contr�le des param�tres impliqu�s.
����������� Le champ �lectrique est constitu� des charges
spatiales dans l'atmosph�re. Les porteurs de ces charges spatiales peuvent
�tre des particules en suspension ou des composants des gaz atmosph�riques.
Lorsque le ciel est clair, les charges spatiales positives pr�dominent et
sont partout de la m�me valeur que l'ensemble des charges n�gatives de la
surface de la terre. C'est pourquoi par beau temps, le vecteur d'intensit�
de champ est dirig� vers la terre (le gradient de potentiel est d'environ
100 V/m). Alors que la diff�rence de potentiels entre terre et atmosph�re
augmente avec l'altitude, le gradient de potentiel d�cro�t.
����������� Dans des zones de mauvais temps ou m�me pendant une
p�riode nuageuse, les conditions sont diff�rentes. Non seulement le gradient
de potentiel peut alors atteindre des valeurs tr�s �lev�es (valeur absolue
maximum mesur�e: 11.000 V/m en �t� 1977) et pr�senter de consid�rables variations
dans le temps, mais il peut �galement changer de polarit�. Lorsque ceci se
produit, l'intensit� de champ change naturellement de direction et la surface
de la terre peut devenir positive. La valeur de gradient de potentiel ne d�pend
pas seulement du moment de la journ�e et de la saison, mais aussi et surtout
de l'altitude et du type de terrain; les valeurs obtenues dans les cavit�s
sont plus faibles et celles obtenues au dessus des arbres, des montagnes etc.
Elles sont plus �lev�es, comparativement � celles relev�es sur des zones planes.
Les champs �lectriques, y inclus ceux �manant des organismes vivants sont
mesur�s, soit en utilisant une sonde de potentiel ayant subi un traitement
radioactif gr�ce auquel le potentiel du point de l'espace de la sonde est
mesur�, soit en utilisant des moulins de champ qui enregistrent les charges
d'induction �lectrostatiques du champ. Il faut souligner que les valeurs obtenues
via ces tests ne repr�sentent pas des valeurs exactes, mais que celles-ci
d�pendent des propri�t�s physiques des indicateurs, comme la capacitance d'entr�e,
la r�sistance d'entr�e et les constantes de temps.
����������� Les courants �lectriques atmosph�riques sont engendr�s
par le mouvement des charges spatiales constitu�es des ions atmosph�riques.
De petits ions sont cr��s sous l'influence des rayons cosmiques et de la radioactivit�
terrestre. Ils d�terminent � la fois la conductivit� atmosph�rique et la compensation
de potentiels entre les corps charg�s. La quantit� d'ions par unit� de volume
d'air augmente avec l'�l�vation en altitude, comme cela a lieu lors de l'�clatement
d'un orage. La combinaison r�sultante de petits ions avec des particules en
suspension neutres � l'origine, aboutit � la formation de gros ions � mobilit�
tr�s r�duite. Ce processus est renforc� par la pr�sence de brume ou de brouillard
ainsi qu'� proximit� des villes et des zones industrielles.
Animaux en vol
�����������
Les oiseaux construisent habituellement leur
nid dans les creux des arbres ou sur une branche sous un d�me de feuillage.
Dans les deux cas les nids sont galvaniquement connect�s � la terre gr�ce
� la conductivit� �lectrique de l'arbre et ils sont presqu'enti�rement
prot�g�s par l'�cran de verdure contre les champs �lectrostatiques atmosph�riques.
La r�sistivit� sp�cifique moyenne de l'atmosph�re est 1015
W/cm et est ainsi oppos�e � celle
des plantes ou � celle d'un sol relativement sec avec des valeurs approximatives
de 105 W/cm. C'est pourquoi
les plantes, comparativement avec la haute atmosph�re fortement isolante,
agissent comme des conducteurs par lesquels le champ �lectrique atmosph�rique
sera continuellement d�charg�.
����������� Cependant, lorsque les oiseaux volent
librement dans l'air, ils sont confront�s avec des courants �lectriques
et des forces �lectriques. La m�me chose s'applique aux insectes, lorsque
par exemple, ils s'envolent d'une surface vers l'air libre. Aucun de ces
tests n'a �t� r�alis� jusqu'ici sur des chauve-souris mais on peut supposer
qu'elles sont sujettes � des processus similaires. En vol, le corps de
l'animal se charge �lectrostatiquement ou s'il est d�j� charg�, il se
d�chargera et sera en m�me temps polaris� di�lectriquement. Cette polarisation
est caus�e par le champ du voltage atmosph�rique direct d�pla�ant les
charges du corps de l'animal � un point tel qu'un dip�le puisse �tre �ventuellement
induit. Le moment de ce dip�le est dirig� dans la direction du champ de
l'excitateur �lectrique; en d'autres mots, verticalement. Le potentiel
de l'espace environnant d'un animal est �quivalent � celui du champ de
polarisation, plus une composante de perturbation engendr�e par le dip�le
de l'animal. Si nous imaginons qu'au lieu de voler, l'animal est plac�
sans mouvement dans l'atmosph�re ionis�e, alors le potentiel environnant
d�terminerait la puissance et la rapidit� du flux des ions atmosph�riques
unipolaires vers lui. Des compensations de charges appara�traient� en
proportion avec la densit� ionique au moyen de courants de conduction
avec une constante de temps d�finie. Des charges spatiales s'accumuleraient
autour du corps et ainsi donneraient une impression ext�rieure de neutralit�
�lectrique.
����������� La compensation de charges ne peut pas se produire
de la m�me mani�re durant le vol pour les raisons suivantes: lorsqu'un
animal vole, l'air circule de fa�on laminaire autour de son corps, ce
qui signifie que ce n'est que la couche tr�s interne d'air qui sera capable
d'induire une compensation de potentiel. Cependant, la concentration de
charges est trop faible. Ce courant de flux d'air laminaire ne s'applique
pas aux ailes, car le courant se dissipe dans les turbulences � la surface
sup�rieure des ailes. En pr�sence de courants de vents en rafales, lesquels
engendrent� une distribution statistique de charges dans l'atmosph�re,
des courants de convection permettent � une compensation entre la charge
de l'aile et l'air environnant, de se mettre en place tr�s rapidement.
De plus, �tant donn� les vitesses des turbulences, les ailes peuvent alors
recevoir une autre charge, dont la polarit� pourrait �tre oppos�e. Cependant,
cet effet est insignifiant, comparativement aux processus enregistr�s
sur des ailes en train de battre. Dans ce cas, il y a avant tout une augmentation
de ce que l'on d�signe par "�lectricit� de friction", par laquelle
les plumes des ailes montrent une pr�f�rence pour les charges positives
correspondant � leurs constantes di�lectriques relatives. Deuxi�mement,
l'activit� de battement chasse toutes les formes de porteurs de charges
du volume d'air concern�. La charge de l'aile et la polarit� de l'aile
correspondent � la densit� de charge spatiale des couches d'air travers�es.
����������� Tous ces ph�nom�nes �lectriques pr�sent�s ici
en termes quelque peu simplifi�s aboutissent au profil de distribution
suivant de polarit� et de potentiel chez les animaux volants (fig.
3). Le corps forme un dip�le vertical induit qui, cependant
ne peut �tre cons�quent que si l'effet du champ �lectrique atmosph�rique
n'est pas affaibli de mani�re d�cisive par la position des points d'inversion
sup�rieur et inf�rieur des ailes en cours de battement. Les extr�mit�s
des ailes, en particulier, sont charg�es durant le battement, en corr�lation
avec la densit� de charges de l'espace environnant. Par beau temps, la
charge est positive. Etant donn� les propri�t�s isolantes �lectriques
�lev�es des plumes (ou de la cuticule chez les insectes), les charges
capt�es sont� pi�g�es � leur point d'impact et deviennent ainsi des g�n�rateurs
de leur propres champs �lectriques. Les param�tres sp�cifiques aux animaux
d'intensit� de champ sont la fr�quence des battements d'ailes et la superficie
de l'aile en contact avec les turbulences de l'air environnant.
����������� Des tests sur des pigeons ainsi que sur des insectes
volant en libert�, r�alis�es dans des tunnels a�rodynamiques, ouverts
ou ferm�s, � la fois � l'air libre et en laboratoire, ont indiqu� que
les tunnels a�rodynamiques en m�tal ne correspondent pas aux conditions
physiologiques naturelles. L'intensit� des valeurs de champ �manant des
animaux dans les tunnels �tait r�duite quasiment � z�ro dans les tunnels
a�rodynamiques alors que par beau temps � l'air libre, des valeurs minimales
comprises entre 10 V/m et 100 V/m �taient r�guli�rement observ�es. Des
tests ioniques, r�alis�s sur les courants d'air laminaires des tunnels
ont montr� que les petits ions qui �taient absorb�s dans le tunnel � partir
de la chambre, se d�chargeaient sur les treillis de m�tal raccord� � la
terre ainsi que sur les parois et n'atteignaient jamais r�ellement les
animaux volants. Ce n'est que lorsque le courant d'air a �t� ionis� au
moyen de deux g�n�rateurs b�ta (tritium) et le tunnel reconstruit presqu'enti�rement
en bois que des valeurs comparables ont �t� obtenues.
|
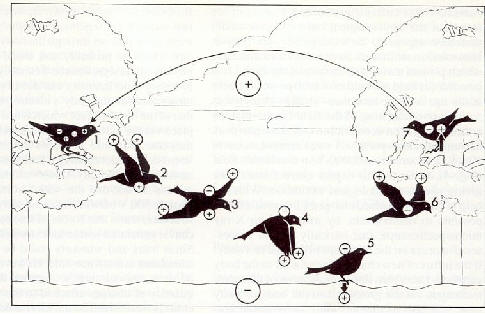
Fig. 3: Charge �lectrique et polarisation d'un oiseau (analogue � un insecte volant) au moyen d'�lectricit� de contact et induction d'un dip�le, se formant dans le champ du condensateur �lectrique atmosph�rique naturel contenant les charges spatiales. Lorsque le contact galvanique avec la terre a lieu (via un arbre, une maison etc.), les charges libres du corps sont neutralis�es (oiseaux 1, 5, 7); les d�mes de feuillage font l'effet d'une cage de Faraday et l'effet de dip�le n'a plus lieu (oiseaux 1, 7). De grandes quantit�s de charges �lectrostatiques sont produites dans un espace d'air �lectriquement isolant (oiseaux 2, 3, 4, 6). Avec tous nos remerciements au Prof. E. Morgenstern, Institut de Microscopie �lectronique, de l'Universit� de la Saar pour pouvoir utiliser cette figure. |
Un capteur de signaux fix� sur un avion enregistre le vol d'un animal comme un champ �lectrique alternatif dont la fr�quence co�ncide avec celle des battements d'ailes. L'intensit� du champ diminue vers l'ext�rieur en proportion inverse du carr� de la distance du bord d'attaque de l'aile (fig. 15).
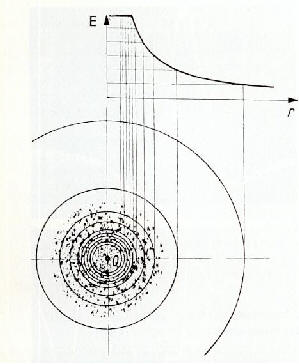
����������� Le corps d'un animal expos� au champ �lectrique sur le sol sera influenc� comme d�cit ci-dessus dans le cas des animaux en vol. Si l'influence provient de charges positives, les charges n�gatives de l'animal seront li�es. Les contre-charges positives qui auparavant �taient coupl�es avec les charges n�gatives et rendaient ainsi le corps neutre, sont donc libres. Pendant un certain laps de temps, elles peuvent �tre compens�es par des �lectrons du sol via les membres. Pendant ce temps les ions positifs correspondant au gradient de potentiel fluent continuellement vers l'animal sous forme d'un courant conduction. Ces courants sont inf�rieurs aux courants physiologiques � raison de plusieurs puissances de dix et de ce fait semblent �tre biologiquement inactifs. Cependant, la plupart des insectes ont des antennes innerv�es qui sont en saillie au dessus de la surface du corps. Il est possible que des intensit�s de telles puissances se d�veloppent aux pointes de ces antennes. Ces intensit�s sont d�peintes visuellement comme un faisceau intense des lignes de champ. L'acc�l�ration rapide des ions qui s'ensuit va engendrer des pics de courants en ces points. Une valeur totale de courant de 10-8 Amp�res mesur�e dans un champ de condensateur de 5000 V/m (fig.4) conduit � s'attendre � des intensit�s de courant donnant lieu � une excitation physiologique. A l'aide d'a�rosols charg�s de CsCl (chlorure de caesium) et finement vaporis�s, il a �t� possible de d�montrer � l'aide d'un microspectroscope � rayons, X que des processus semblables se produisent sur des micropointes en saillie du corps de l'insecte. Si l'influx des nouvelles charges de l'air vers la surface du corps est plus rapide que la neutralisation des contre-charges, via le sol, alors, le corps ou des parties du corps prendront des charges jusqu'� ce que le gradient de potentiel par rapport � l'environnement atteigne z�ro. Le temps mis pour qu'une compensation compl�te de potentiel ait lieu est en corr�lation avec les constantes de l'atmosph�re au cours du temps, au niveau du sol et �galement avec la zone superficielle impliqu�e de l'animal (souris : t >30). Si, par exemple, un lapin se glisse sous le sol dans son terrier alors qu'il est charg� de cette mani�re, il devient ainsi "blind�" contre le champ �lectrique atmosph�rique ext�rieur qui avait auparavant agi comme facteur neutralisant. La fourrure de l'animal charg�e du flux d'ions engendre maintenant son propre champ �lectrique.
 |
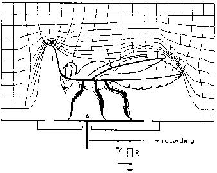 |
����������� Fig.4 : Relations di�lectriques chez des insectes dans le champ du condensateur naturel. Les parties expos�es du corps (antennes, bouts des ailes, micro-saillies) provoquent une concentration de lignes de champs et une conduction de pics de courant. Les zones du corps ayant une r�sistance ohmique relativement basse par rapport � la terre (membranes) ont des effets similaires. Des densit�s de courant physiologiquement actives peuvent se produire dans les organes de perception situ�s dans ces zones. Des poils sensitifs peuvent �tre stimul�s m�caniquement dans des r�gions contigu�s � des structures � haute r�sistance ohmique (scl�rites), � cause des augmentations d'intensit�s de champ (voir aussi la fig. 22). Le dessin des lignes de champ a �t� trac� � partir d'un insecte qui a �t� plac� comme �lectrode n�gative dans un conteneur rempli de t�trachlorure de carbone dans lequel de fins filaments d'ac�tate �taient suspendus. Lorsque la tension (voltage) a �t� appliqu�, les filaments se sont organis�s le long des lignes de champ. (R = r�sistance de mesure).�
Ceci a deux conséquences distinctes. Tout d'abord, un courant de déplacement s'écoule à travers le corps avec une intensité qui est probablement trop faible pour avoir une quelconque signification physiologique. Deuxièmement, une intensité de courant extrêmement forte peut se développer à une distance très proche entre le système d'électrolytes du corps (par exemple celui de la surface nasale qui, lorsque la mise à la terre a lieu via les pattes, a virtuellement le potentiel zéro) et les poils de la fourrure. L'intensité du champ est une fonction où prédomine l'humidité atmosphérique relative. Des observations réalisées sur une surface plane de 1 dm² avec une tension (voltage) d'environ 500 Volts entre le système d'électrolytes et la structure de la fourrure montre que des forces de l'ordre de grandeur du kilopoids peuvent être engendrées dans un espace extrêmement restreint. Des poils des sinus et des vibrisses (moustaches) peuvent être stimulés mécaniquement dans ce champ de forces. Cependant, les terriers de la plupart des animaux sont si étroits que la plus importante quantité des charges accrochées à la fourrure est presqu'entièrement neutralisée par l'inévitable contact avec la terre.
Une attention particulière doit
être accordée aux insectes. La surface du corps de presque tous
les insectes n'est pas seulement composée de sclérites à
haute résistance qui accumulent les charges comme décrit ci-avant,
mais aussi de structures membranaires à relativement faible résistance
électrique qui peuvent être mises à la terre via le système
d'électrolytes (hémolymphe) et des membres (sécrétions
adhésives). C'est pourquoi on peut s'attendre à trouver des
forces dans l'espace situé entre les sclérites et les membranes,
capables de stimuler les poils sensitifs mécaniquement hautement sensibles.
Pi�zo-�lectricit�, pyro-�lectricit� et ph�nom�nes d'�lectret
��������������� L'�lectricit� peut �tre non seulement engendr�e par induction �lectrostatique, par polarisation et par ce que l'on appelle des processus de friction ou de contact, mais aussi par d'autres m�canismes importants chez l'animal comme l'effet pi�zo-�lectrique et l'effet pyro-�lectrique. La r�p�titivit� de r�sultats exp�rimentaux obtenus est si importante qu'elle permet d'attribuer une signification cl� � ces effets. Dans beaucoup de tissus d'organes des t�guments des vert�br�s et des arthropodes, comme les structures de k�ratine, de collag�ne, de cellulose et de chitine, des charges n�gatives sont accumul�es lorsque les tissus se tordent ou sont sujets � une pression ou sont �tir�s dans une direction particuli�re. L'extr�mit� de la surface de l'�chantillon pr�sente alors des charges positives [3, 4, 8, 9, 13, 18]
����������� Une alternance entre compression et dilatation
provoque un changement de signe de la charge et la valeur de la charge
m�canique est proportionnelle � la production de charges �lectriques.
Tous les tissus dans lesquels de la piezo-�lectricit� est observ�e sont
�galement � effet pyro-�lectrique, gr�ce auquel le r�chauffement et le
refroidissement induisent la production de charges avec inversion ad�quate
de signe. Ces effets sont probablement caus�s par des moments dipolaires
permanents qui sont compens�s � l'ext�rieur par des charges superficielles
lorsque le tissu est au repos. C'est seulement lorsque le moment �lectrique
mol�culaire des structures du tissu est modifi� par une influence m�canique,
ou lorsqu'un changement de temp�rature modifie la charge superficielle
que l'aptitude � compenser totalement l'effet �lectrique n'est plus possible.
 |
���������������
Fig.5 : Image d'une �mission photo-�lectronique de la structure
de tissu d'une coupe transversale de cuticule. Les �lectrons sont �galement
�mis par la cuticule de l'insecte dans la lumi�re solaire. Ce processus
modifie le potentiel superficiel du corps. Stimulant : lumi�re polychromatique
HBO 100; grossissement 1250 x.
Avec nos remerciements au Prof. Dr. E. Morgenstern, Institut
de Cytologie et de Microscopie �lectronique (Univ. de la Sarre) pour la
pr�paration de la coupe et au Dr. Engel du Fritz Haber Institut � Berlin
pour la r�alisation de l'image.
Les
valeurs de test correspondent aux exc�s sp�cifiques de charges. Cette
forme de production de charge �lectrique est �galement particuli�rement
�vidente en thermo-�lectricit�, en photo-�lectricit� et pour les r�cepteurs
m�caniques. En somme, il est probable que dans les diff�rentes formes
de vie, un m�canisme pyro-�lectrique et un m�canisme pi�zo-�lectrique
soient impliqu�s dans la perception de la chaleur, de la lumi�re et des
vibrations [3].
Beaucoup d'épithéliums biologiques sont équipés
d'une couche de cire protectrice. Cette couche protectrice peut également
avoir la structure d'un dipôle électrique permanent répondant
aux lois de la physique de l'électret. La cire d'abeilles liquide,
exposée à un champ électrique se solidifiera également
en un électret 2
[28].
L'interaction entre lumi�re et �lectricit�
est vari�e dans la mati�re organique. Une conductivit� �lectrique augment�e
est observ�e dans tous les tissus biologiques examin�s jusqu'ici (insectes,
oiseaux, mammif�res) lorsque des photons d'�nergie suffisante sont absorb�s
(effet photo-�lectrique interne). La description de l'effet photonique externe
semblerait mieux r�pondre au th�me du pr�sent document : un ph�nom�ne qui
a jusqu'ici fait l'objet d'�tudes plus pouss�es pour les insectes. C'est pourquoi
nous donnerons ci-apr�s une br�ve description qualitative de deux exp�riences
simples mais concluantes.
a) Un insecte (aussi frais que possible) est placé dans un conteneur
en verre rempli de tétrachlorure de carbone (CCl4). Des filaments
d'acétate de sodium sont suspendus dans le tétrachlorure
de carbone. Si le champ d'un condensateur est mis en place de telle manière
que l'insecte fonctionne comme une électrode négative, les
filaments d'acétate de sodium s'arrangent selon les lignes de flux
électrique et se terminent à la surface du corps de l'insecte
(voir figure 4).
La cuticule de l'insecte peut être exposée à des petits
points d'énergie lumineuse en utilisant une fine fibre conductrice
de la lumière. La photoréaction en ces points consiste en
une dispersion explosive localisée des filaments d'acétate.
Le même effet peut être obtenu en utilisant un laser.
b) Un tissu de cuticule est desséché à froid (lyophilisé)
et pulvérisé dans un mortier. Quelques particules sont insufflées
dans un champ capacitif vertical et stabilisées dans un état
de suspension [34].
Si les particules stationnaires sont alors irradiées par de la
lumière ultra-violette, elles se dirigent vers la plaque négative
du condensateur.
On peut conclure à partir de ces deux expériences que des
électrons sont émis par la cuticule de l'insecte sous l'influence
de la lumière (figure
5). Le processus quantitatif de ce phénomène
est hautement complexe.
Implication biologique des champs �lectriques
Dans la solution de certains probl�mes d�finis du domaine de la biologie, il s'est r�v�l� judicieux d'explorer et de d�couvrir les m�canismes du syst�me concern� par l'observation. Ceci a conduit dans la suite � la confirmation du principe selon lequel il existe une quantit� de bio�nergie qui peut �tre capt�e et utilis�e de multiples mani�res par les animaux. Ainsi, si l'on explore l'implication d'une charge �lectrostatique de la surface du corps, un processus comprenant une vari�t� de composantes physiques, deux effets connus depuis quelques temps d�j� peuvent appara�tre chez les oiseaux, les mammif�res � fourrure et quelques insectes.
Dans le cas des insectes
pollinisateurs, le rendement en pollen et la distribution de celui-ci sont
augmentés par les forces d'attraction entre la surface du corps chargée
positivement et le pollen généralement chargé négativement.
La régulation thermique chez les oiseaux et les mammifères est
rendue indirectement optimale par les forces électrostatiques. Ceci
se produit grâce aux plumes du duvet qui sont individuellement chargées
négativement parce qu'elles frottent contre le plumage extérieur
de couverture. Etant ainsi de même polarité, elles se repoussent
mutuellement. Elles remplissent ainsi si complètement la couche que
la convection de l'air est virtuellement mise à l'arrêt et la
chaleur du corps est maintenue. Cet effet est renforcé par le fait
que les plumes externes chargées du signe opposé (c'est-à-dire
positivement) sont attirées contre la couche interne diffuse en raison
des lois de l'électrostatique. Nous sommes donc capables de trouver
plusieurs effets biologiques utiles comprenant quelques cas qui n'échappent
pas jusqu'ici à cette hypothèse.
Les résultats de nos expérimentations sur l'influence des
facteurs électriques exogènes sur le comportement animal nous
ont fourni des lignes directrices. Il n'y a pas de doute que toutes les composantes
électriques de la surface de l'animal décrites ici peuvent être
altérées par des facteurs électriques artificiels et
naturels en ayant une répercussion sur des effets physiologiques.
En adjoignant la littérature sérieuse concernant ce sujet, l'influence
de paramètres physiques peut être classée comme suit,
selon les effets biologiques qui en résultent:
1. Selon la fréquence et le type
de champ: champ électrique alternatif, champ magnétique alternatif,
oscillations électromagnétiques (à extrêmement
basses fréquences -ELF-, à très basses fréquences
-VLF-).
2. Selon le contenu d'énergie:
intensités de champs beaucoup plus puissantes que celles qui se manifestent
dans l'environnement naturel, intensités de champ du même ordre
de grandeur que celles rencontrées dans la nature.
Dans un laboratoire, les oscillations
électromagnétiques ne peuvent être appliquées qu'au
dessus d'une plage relativement basse de longueurs d'ondes. En effet, les
expériences réalisées avec des oscillations à
très grandes longueurs d'ondes, donc à très basses fréquences
(de 3 Hz à 30 kHz) ne peuvent être prises en considération
pour démontrer l'influence d'un champ électromagnétique.
Dans cette plage de fréquences, les effets sont produits, soit par
les champs magnétiques alternatifs émis par une bobine, soit
par les champs électriques alternatifs engendrés entre deux
plaques de métal. Dans le cas du champ d'un condensateur, il est nécessaire
de vérifier dans chaque cas, si la charge capacitive dans les fils
d'alimentation électrique n'a pas été la cause du passage
d'un courant, lequel pourrait avoir produit des composantes de champs magnétiques
ayant une action biologique additionnelle. A l'air libre, d'autre part, il
existe un risque de confusion entre les oscillations électromagnétiques
de la plage des basses fréquences découvertes, car leur présence
peut être en corrélation avec des valeurs produites par des phénomènes
météorologiques.
En tenant compte des conditions décrites ci-dessus, nous avons découvert
que l'influence de paramètres électriques induisent les effets
suivants.
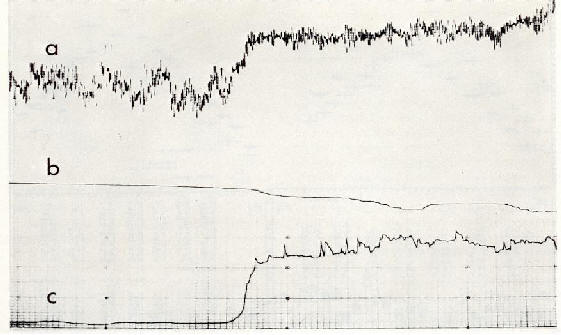
����������� L'activit� motrice de souris blanches
est rapidement augment�e lors de l'exposition � un puissant champ �lectrique
alternatif 50 Hz (5 kV/m). Les animaux s'adaptent individuellement � ce
stimulus de mani�re diff�rente, apr�s des p�riodes s'�tendant de quelques
heures � plusieurs jours. Des animaux de race g�n�tiquement identique,
accoutum�s aux conditions de laboratoire, ont montr� des variations analogues
au niveau sanguin, c'est-�-dire une augmentation du taux de gamma-globulines
et une diminution des leucocytes suite � une exposition � un champ �lectrique
(40 kHz; 10 V/m) comparativement � des souris prot�g�es par �crans. Les
modifications des figures sanguines d�pendent de la dur�e de l'exp�rimentation.
Des animaux examin�s apr�s 7 et 14 jours d'exp�rimentation ne diff�raient
pas des sujets t�moins (Fig.6)[2].
Ces recherches et les r�sultats de l'�valuation du taux de corticost�rone
du cortex surr�nal de ces m�mes animaux [20]
indiquent que le champ �lectrique alternatif est un stimulus de stress.
Ce ph�nom�ne peut �tre plus clairement per�u chez des insectes vivant
en colonies. L'effet des champs �lectriques 50 Hz sur le comportement
des colonies d'abeilles a �t� explor� dans des s�ries d'exp�rimentations
en laboratoire [39-40]
et corrobor� par un film. Les abeilles deviennent tr�s agit�es � approximativement
11.000 Volts/m et la temp�rature de la ruche s'�l�ve fortement. La d�fense
du territoire s'intensifie � tel point que des abeilles de la m�me colonie
s'attaquent entre elles. Apr�s quelques jours sous l'influence du champ
�lectrique, les abeilles extraient le couvain des alv�oles et n'en replacent
pas de nouveau. De plus, le miel et le pollen sont consomm�s et plus aucune
r�colte n'a lieu. Les abeilles qui avaient �t� plac�es dans la ruche peu
de temps avant le d�but de l'exp�rience ont r�guli�rement quitt� la ruche
pendant la p�riode d'application des champs �lectriques.
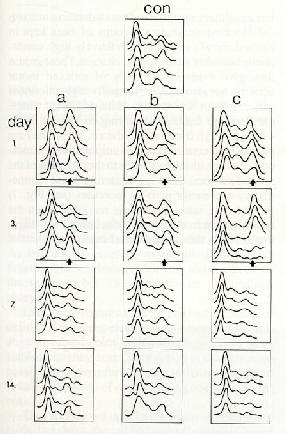 |
����������� Fig.6 : Enregistrement de la s�paration par �lectrophor�se des prot�ines
du s�rum de souris apr�s diff�rentes dur�es d'exposition � un puissant
champ �lectrique, comparativement au s�rum de souris prot�g�es par une
cage de Faraday:
������������������con
= t�moins (exposition simul�e)��
������������������a) champ alternatif 40 kHz,
� environ 5 kV/m
������������������b)
champ continu 5 kV/m
������������������c) �cran de blindage contre
les champs �lectriques (cage de Faraday)
L'�norme augmentation de gamma-globulines (fl�ches) enregistr�
durant les premiers jours indique l'influence du facteur stress, � la
fois en pr�sence et en l'absence du champ �lectrique. L'adaptation est
possible (4-14 jours) [2].
�����������
D'autre part, les abeilles qui �taient d�j�
bien accoutum�es � leur ruche avant l'exp�rience ont commenc� � sceller
toutes les crevasses et les trous, y compris le trou de vol, avec de la
propolis. Le manque d'oxyg�ne a conduit � une ventilation accrue, causant
une �l�vation aigu� de la temp�rature et la mort �ventuelle de la colonie
[45].
����������� Des colonies tr�s
sensibles ont d�j� pr�sent� des signes mesurables de r�actions lorsqu'elles
ont �t� expos�es � des intensit�s de champ �lectrique d'environ 100 V/m �
des fr�quences comprises entre 30 Hz et 40 kHz. On a pu observer qu'� l'instant
o� le champ �tait enclench�, toutes les ailes se mettaient soudain � battre
� un rythme de 100-150 Hz.
On a pu observer que les oiseaux ne perchent g�n�ralement
pas sur les c�bles de phase des lignes � haute tension, mais sur le fil de
terre.
����������� Etant
donn� que les abeilles d'une m�me colonie, soumises � un champ �lectrique
alternatif ne sont plus aptes � se reconna�tre entre elles, il est possible
que leurs propres canaux d'informations soient endommag�s. Cette supposition
est corrobor�e par le fait que les abeilles sont particuli�rement agressives
lors d'augmentations d'activit�s atmosph�riques dans la plage de 10-20
kHz [37].
Ces jours l�, le taux de retour des abeilles � la ruche est tr�s fortement
r�duit, m�me lorsque le temps est compl�tement calme [5].
����������� Le m�me ph�nom�ne appara�t pour certaines esp�ces
d'oiseaux. Les pigeons rentrant au pigeonnier sont fortement d�sorient�s
pendant des p�riodes d'augmentation de l'activit� atmosph�rique [10]
et il peut se faire que les oiseaux s'affolent dans des plages particuli�res
d'intensit� d'antennes d'�missions radio [11].
Des exp�rimentations syst�matiques ont �t� men�es relativement � la perception
des champs 50 Hz sur des pigeons rentrant au pigeonnier [36].
Les pigeons �taient l�ch�s pr�s de lignes a�riennes � haute tension de
380 kV, de mani�re telle que les oiseaux devaient voler � travers les
champs �lectriques des lignes pour retourner au pigeonnier. Le r�sultat
�tait que chaque pigeon volait soit par dessus le c�ble le plus �lev�
(c�ble de terre), soit sous l'ensemble des c�bles, presqu'au niveau du
sol.. Dans chaque cas, ces choix de ligne de vol se faisaient en fonction
de la derni�re intensit� de champ �lectrique subie. Dans le cas d'une
ligne � haute tension de 110 kV, quelques pigeons volaient �galement entre
les c�bles. On a observ� dans la suite que les vol�es d'oiseaux migrateurs
se scindent r�p�titivement avant de survoler une aire de transformation
(sous-station) et effectuent un d�tour comme pour contourner un obstacle
invisible pour reconstituer ensuite la vol�e.
Variations
m�t�orologiques
����������� Quelques insectes et sp�cialement les abeilles sont r�put�es pour �tre sensibles aux changements m�t�orologiques. Les changements de temps et les passages nuageux sont tr�s �troitement en corr�lation avec les variations de potentiels du champ �lectrostatique naturel. Ces variations de potentiels sont la cause de modifications de potentiels locaux � la fois sur les animaux � titre individuel et sur l'ensemble de colonies d'abeilles [14, 37]. Etant donn� que les insectes sont capables de capter les diff�rences de potentiels dans l'environnement naturel [12, 25, 29, 33, 35], il est hautement probable que l'information concernant la dynamique d'�volution m�t�orologique soit per�ue de cette mani�re (fig 7 et fig.8).
����������� Les niveaux pr�dominants d'humidit� et de temp�rature de l'atmosph�re environnante peuvent �galement �tre per�us simplement par l'insecte parce que la valeur du courant �lectrique de compensation entre des zones de diff�rentes intensit�s de charge sur la cuticule est modul�e de mani�re d�cisive par l'ensemble de ces param�tres physiques.
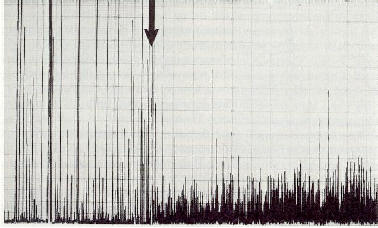 |
�������� Fig.
7 : Battements d'ailes anormaux parmi un groupe d'abeilles dans une cage
de Faraday (pics d'enregistrement �tir�s). Le battement d'ailes est associ�
� une auto-�lectrification et cesse aussit�t que la cage de Faraday est
aliment�es avec des ions atmosph�riques (fl�che). Des ions atmosph�riques
monopolaires induisent une charge �lectrostatique. L'activit� motrice
augmente dans l'atmosph�re charg�e d'ions (pics d'enregistrement courts
et serr�s). [1,
36].
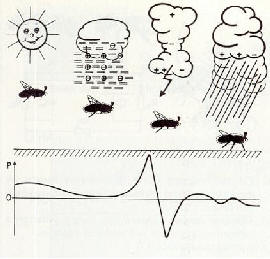 |
���������� Fig. 8 : Potentiel de charge (P de la courbe inf�rieure) d'insectes en vol et d'oiseaux durant des variations m�t�orologiques. La charge d'une colonie d'abeilles se modifie �galement dans le m�me laps de temps.
Les r�f�rences de temps
����������� Les rythmes circadiens d'activit� des colonies d'abeilles ont quelques particularit�s surprenantes. M�me dans des conditions de total isolement (blindage �lectrique), c'est-�-dire dans des conditions constantes, des cycles de fluctuations uniformes apparaissent dans plusieurs groupes non isol�s, aussi bien que chez les groupes sous isolement [36]. L'influence� aussi bien de fortes concentrations d'ions atmosph�riques monopolaires (fig. 9) et de champs �lectriques alternatifs de faible intensit� (1 V/m, 180 Hz) provoquent �galement un d�calage dans le temps des rythmes. Ces d�calages se produisent sur une p�riode de trois jours comparativement � un groupe t�moin non soumis au test. Il est donc possible que certains champs �lectriques fonctionnent comme r�f�rence de temps pour les insectes.
Il est absolument essentiel pour les abeilles d'avoir une charge �lectrostatique individuelle. Des groupes d'abeilles maintenues dans des cages m�talliques reli�es � la terre et soumises � une humidit� atmosph�rique relativement �lev�e vont perdre cette charge.
Ces groupes pr�sentent alors non seulement une diminution de l'activit� motrice, mais aussi une activit� accrue anormale, concert�e de ventilation. En battant des ailes, les abeilles essaient continuellement de se charger par le frottement avec l'air. Lorsqu'elles subissent une atmosph�re � forte concentration en ions unipolaires, qui finalement conduit � la charge individuelle des abeilles, les battements d'ailes cessent imm�diatement et l'activit� motrice augmente (voir fig.7) [1, 36]. La m�me r�action est enregistr�e lorsque les abeilles sont maintenues dans des cages isol�es de PVC au lieu de cages m�talliques reli�es � la terre.
Les oiseaux peuvent �galement �tre observ�s en train de frotter intensivement leur bec contre leurs plumes apr�s un bain. Etant donn� que le bain implique une d�charge extensive, la fonction de frottement n'est pas seulement destin�e � lisser les plumes, mais aussi � r�tablir une charge �lectrostatique.
Tout corps charg� �lectrostatiquement produit un champ �lectrique alternatif lorsqu'il se meut par rapport � un point fixe. Si ce champ alternatif poss�de une fr�quence stable pendant une longue p�riode de temps, alors la possibilit� d'un transfert d'informations vers d'autres individus via un code �lectrique ne peut �tre �cart�e.
Ceci s'applique non seulement aux vols d'insectes et d'oiseaux, mais aussi aux battements de queue et d'ailes dans les communaut�s d'insectes ainsi qu'aux mouvements
����������� Fig.9 : Rythme circadien d'activit� d'un groupe d'abeilles durant des p�riodes
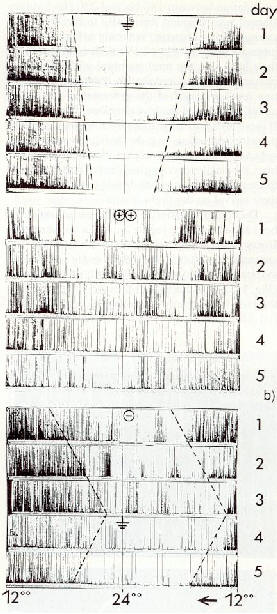 |
de 5 jours sous des conditions appel�es "constantes".
Enregistrements typiques
����������� a)
sous des conditions d'isolement �lectrique : les rythmes tendent � dispara�tre
����������� b)
dans un air fortement ionis� + ou � (approximativement 105
ions /cm�): la phase d'activit� est incoh�rente, sans r�f�rence de temps.
����������� c)
dans un air faiblement ionis� ( - approximativement 103 ions
/cm�): d�calage de la phase d'activit�, r�versible pendant les 3 premiers
jours.
Une remise en place du blindage d�cale la phase dans
la direction oppos�e. Le d�calage de la phase d'activit� dans l'air �
forte concentration en ions implique sa fonction de r�f�rence de temps.
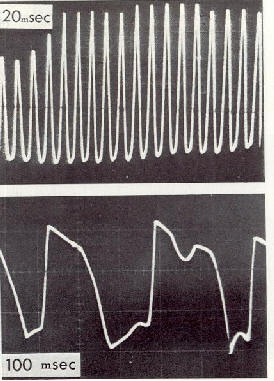 |
���Fig.
10 : Oscillogramme du champ �lectrique alternatif produit par les battements
d'ailes
a) d'une abeille,
b) d'un pigeon dans un tunnel a�rodynamique. �
rythm�s de parties
du corps de l'animal comme par exemple dans toutes les formes de danse
et de contacts mutuels d'antennes en vibration (fig.
10 et fig.
11). Il est physiquement parlant, in�vitable que l'influence
ou l'induction prennent place entre individus proches. Il n'est pas difficile
de prouver que ces champs sont r�ellement assimil�s et trait�s comme des
signaux. Eskov et al. [15,
16] ont �t� les premiers � d�couvrir des r�actions typiques
de r�ponse � des signaux de simulation de danse chez les abeilles. Jusqu'ici,
nous n'avons �t� capables de reproduire que quelques uns de ces r�sultats.
Un message sp�cial semble �tre contenu dans la modulation de fr�quence
entre 180 Hz et 250 Hz. Des abeilles solitaires affam�es r�pondent de
mani�re r�p�titive � ce signal en �levant leurs antennes, mais seulement
lorsque l'intensit� de champ demeure en dessous de 1 V/cm. Les abeilles
approchant de leur ruche modifient le potentiel total de la ruche, de
mani�re caract�ristique et absorbent par contr�le en retour (feed� back),
une partie du potentiel d'un "nuage de charge" devant l'entr�e
de la ruche (fig.
12) [37].
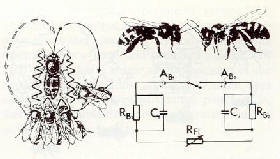 |
Fig.
11 :
a) la danse
des abeilles. La transmission d'informations via des champs �lectriques
alternatifs (modifi�e � partir de von Frisch, 1973)
b) le contact
d'antennes entre deux insectes engendre un courant de compensation et
des impulsions de potentiel [37].
Si ces abeilles sont
charg�es de mani�re "non physiologique" � la fois quant � la
polarit� et quant � la valeur de la charge (par exemple en ayant re�u
de l'eau sucr�e charg�e) avant leur retour � la ruche, elle vont �tre
l'objet d'une grande attention de la part du reste de la colonie, � la
fois avant et apr�s leur arriv�e sur la planche de vol.
����������� La forme communicative du contact par antennes interpos�es
chez les abeilles et les fourmis peut �tre enregistr�e sur un oscilloscope.
Chaque fois qu'un bref contact se produit entre les antennes, un signal
est engendr� dans le syst�me d'�lectrolytes du r�cipient sous forme d'une
impulsion (fig.
11). L'intensit� de l'impulsion est fournie par l'intensit�
de la charge individuelle, la diff�rence de potentiels de contact et par
dessus tout, par les effets capacitifs.
����������� Le signal est cod� en fr�quences. Un courant de compensation
s'�coule lorsque l'antenne ou le corps sont touch�s. L'intensit� de ce
courant est d�termin�e dans chaque cas par la r�sistance �lectrique interne
de l'animal ainsi que par la r�sistance de la surface de parcours. Selon
Galuszka et Lisiecki [19],
diff�rentes colonies d'abeilles ont une r�sistance interne diff�rente,
laquelle rend plausible le fait que les abeilles soient nantie d'une m�thode
leur permettant de distinguer les "amies" et les "ennemies"
gr�ce � la chute de tension (voltage) interne. Les termites semblent �galement
communiquer au moyen de canaux �lectriques; Becker [6]
a trouv� des indications claires de communications mutuelles entre des
groupes isol�s.
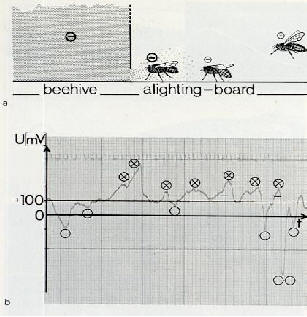 |
Fig.
12 : Relations de potentiel au niveau de la ruche
��������������� a) le potentiel
du corps de chaque abeille volant vers la ruche est modifi� par un nuage
sph�rique de charge qui provient de l'int�rieur de la ruche; il est d�limit�
en caract�ristiques et en dimensions pour chaque colonie.
��������������� b) l'arriv�e
et le d�part de chaque abeille affecte le niveau de potentiel de sommation
de� la ruche. U = abeille
arrivant;� � = abeille partant. [37].
Vol�es d'oiseaux
"Comment une vol�e
d'oiseaux est maintenue ensemble" est depuis longtemps un sujet de
discussions (la m�me question s'applique aux essaims d'abeilles et aux
bancs de poissons). On a observ� en particulier que de tr�s grandes vol�es
d'�tourneaux s'�tendant jusqu'au del� de 500 m� sont capables d'accomplir
des man�uvres tr�s compliqu�es en moins de 5 millisecondes. La question
est: "Comment ces oiseaux peuvent-ils percevoir et ex�cuter le signal
end�ans une fraction de seconde, et ce � quelqu'endroit qu'ils occupent
dans la vol�e?"
����������� Il est bien connu que la transmission de son dans l'air requiert
un temps consid�rable et que le positionnement d'un leader par voie optique
est emp�ch�e par les membres participant � la vol�e. Heppner et Haffner
[22]
ont formul� l'hypoth�se selon laquelle les man�uvres en vol seraient coordonn�es
par des signaux �lectromagn�tiques. De tels signaux ont une rapidit� de
dispersion approchant la vitesse de la lumi�re et pourraient donc atteindre
tous les oiseaux "simultan�ment" quelle que soit leur position
dans la vol�e. Cette hypoth�se gagne � �tre prise en compte, �tant donn�
que les animaux sont charg�s �lectrostatiquement pendant le vol. Il est
possible de d�montrer au moyen d'un oscilloscope que le rassemblement
d'animaux volant individuellement engendre un champ continu r�sultant
(g�n�ralement positif) modul� par les battements d'ailes de tr�s faible
amplitude (fig.
13). Cette modulation doit �tre consid�r�e comme un battement
collectif de tous les battements individuels. La distance entre le r�cepteur
des signaux et l'�metteur particulier est d�cisive. La fr�quence de battement
des oscillations est toujours inf�rieure aux fr�quences des battements
individuels des ailes alors
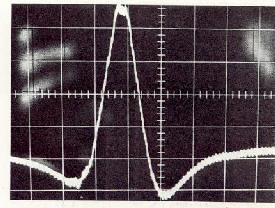 |
que l'amplitude
maximale de battement des oscillations est toujours plus �lev�e que les
amplitudes des battements d'ailes individuels. Les valeurs mesur�es varient
en fonction des conditions m�t�orologiques et des relations g�om�triques
(fig. 14
et fig. 15).
Les donn�es obtenues jusqu'ici indiquent que des vol�es de petits oiseaux
volant � une altitude d'environ 40 m ont un voltage de plus de 6000 Volts.
Jusqu'ici, on ne peut que sp�culer sur la nature des signaux cod�s utilis�s
dans les man�uvres des virages, en postulant que la fr�quence r�elle de
battement et la valeur de l'amplitude de battement sont fix�es pour chaque
oiseau individuellement. Si cette valeur d�cro�t, elle serait imm�diatement
corrig�e par un changement ad�quat de direction de vol.
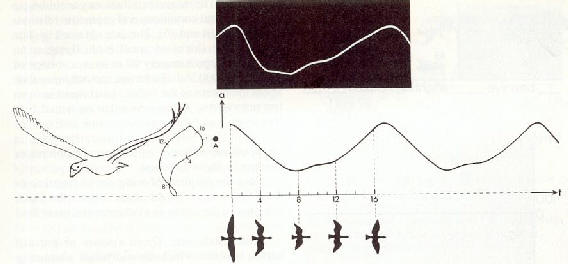 |
�����������
Fig. 14 : Elaboration du champ alternatif
d'un battements d'ailes � partir des rapports de distance entre le trac�
vertical dans l'espace du battement d'ailes et un r�cepteur de signaux.
Au dessus, l'oscillogramme du champ alternatif du battement d'ailes enregistr�
dans un tunnel a�rodynamique � l'aide d'une sonde de potentiel situ�e
au point A. Les trac�s des courbes sont bien parall�les.
����������� Etant donn� que la majorit� des fr�quences de battements d'ailes ont approximativement la m�me valeur, la vol�e appara�t comme un tout en un champ �lectrique coh�rent dans l'espace.
����������� Quelques
petits oiseaux volent en pouss�es d'impulsions entre lesquelles, le champ
alternatif individuel est z�ro. Cette p�riode pourrait �tre utilis�e pour
obtenir un contr�le non perturb� du champ �lectrique de l'oiseau voisin.
 |
Relations
entre les forces �lectrostatiques en vol en formation
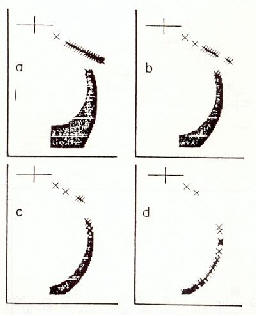
�������� Des esp�ces d'oiseaux qui volent en formation maintiennent un ordre typique m�me lorsque les oiseaux ne sont que deux. Le second oiseau vole � c�t� et en retrait derri�re le premier. Plus l'espacement lat�ral est grand, plus l'espacement en direction du vol est grand [30].
����������� Les relations
de champ �lectrique qui se produisent dans l'atmosph�re sont pr�sent�es
dans un mod�le exp�rimental sur les figures 16 a-c. Les plus fortes intensit�s
de champ se produisent � la r�gion du bec, dans la zone de la queue et
aux extr�mit�s des ailes. L'oiseau volant derri�re re�oit une charge alternative
induite �lectrostatiquement dans la r�gion t�te-cou � partir des ailes
de l'oiseau volant devant. Entre les ailes et la t�te un champ �lectrique
augment� est form�. En m�me temps, le battement d'ailes de l'oiseau positionn�
en arri�re induit des charges �lectrostatiques dans les pattes de l'oiseau
volant devant. Les charges �lectrostatiques induites cr�ent un champ �lectrique
comme indiqu� sur le mod�le. Ainsi, le deuxi�me oiseau est influenc� �lectrostatiquement
par son propre mouvement, et par la pr�sence de l'oiseau volant devant.
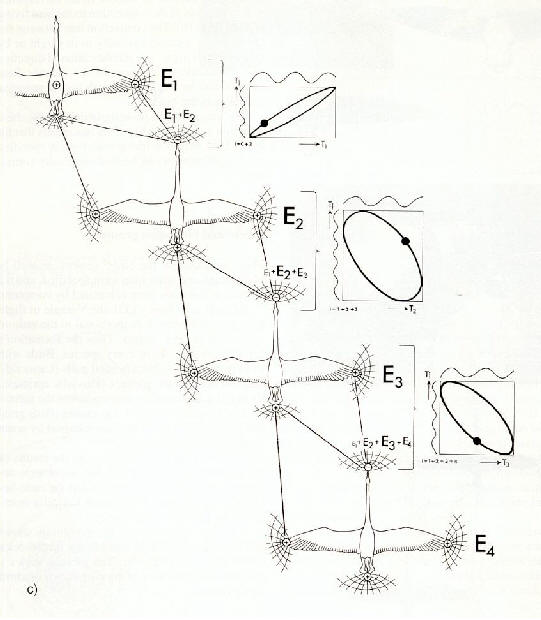
����������� La figure 17 illustre les relations des forces �lectriques
dans une formation compos�e de plusieurs oiseaux. Chaque oiseau est li�
� tout autre oiseau par des champs �lectriques d'intensit� et d'orientation
d�termin�es. Il faut prendre en consid�ration que le corps de l'oiseau
est conducteur en raison des �lectrolytes du corps et que cela suppose
une forme de dip�le dans sa dimension lin�aire. Un champ alternatif superpos�
r�sulte du mouvement rythmique des ailes des oiseaux volant � l'avant.
L'intensit� du champ d�pend de la distance entre les oiseaux et/ou l'envergure
des ailes.
����������� Deux composantes de forces de diff�rentes directions concernent
la r�gion de la t�te de tout oiseau dans la formation (fig.
16). Ceci aboutit � une somme vectorielle de grandeur et de
 |
��������������� a) deux oies en formation de vol
��������������� b) repr�sentation du mod�le exp�rimental de la relation du champ �lectrique et des forces de Coulomb entre deux oies en alignement de formation
��������������� c) distribution de champ entre les oiseaux, montrant un parall�logramme de vecteurs de forces
direction d�finies. L'hypoth�se consiste � dire que la grandeur et la direction de telles forces sont perceptibles et peuvent �tre impliqu�es dans des processus neurophysiologiques. Les oiseaux volent en ligne droite lorsque la direction de la somme des champs �lectriques correspond � la direction de la connexion � l'oiseau volant en t�te (fig. 16). La ligne de connexion entre les t�tes peut �tre localis�e visuellement � la lumi�re du jour ou par des cris la nuit. L'identification de la direction et de la grandeur de la somme des vecteurs de champ �lectrique peut �tre possible via des r�cepteurs tr�s sensibles sur le cou.
����������� L'exigence de congruence de la direction du champ avec la ligne de t�te � t�te signifie que pour chaque oiseau volant en formation, une position sp�cifi�e peut �tre formul�e math�matiquement.
G�om�trie calcul�e de la formation
����������� Les coordonn�es
des centres des oiseaux, ainsi que la longueur de corps et l'envergure
des ailes sont sp�cifi�es et les positions dans la formation sont calcul�es
par computer.
����������� Selon Poncy [32],
l'angle du V de la formation en vol est inversement proportionnel au rapport
entre l'envergure des ailes et la longueur du cou. Donc, la g�om�trie
de la formation est sp�cifique � chaque esp�ce. Les oiseaux poss�dant
un cou court comme les mouettes rieuses (Laris ridibundus) et les pluviers
dor�s (Pluvialis apricaria) volent en formation V � angle applati alors
que les formations d'oiseaux � cou long (par exemple les grues (Grus grus)
et les oies (anseridae) volent en formation V � angle aigu.
����������� Les observations de Poncy sont en accord avec les r�sultats
des calculs faits selon l'hypoth�se d�crite ci-avant. La figure
18 montre que le rapport entre l'envergure des ailes et la
longueur du corps d�termine de mani�re fondamentale la g�om�trie de la
formation.
����������� Une autre application de l'hypoth�se montre que plus la distance
lat�rale est grande, plus l'oiseau doit voler loin derri�re (fig.
19). La figure montre l'optimisation successive de la position
x-y d'un oiseau volant derri�re.
 |
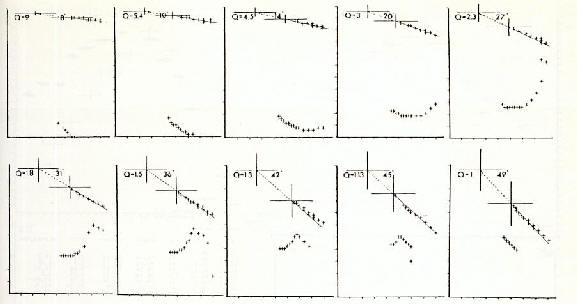 |
V�rification de l'hypoth�se sur base de documents photographiques de formations de vol
����������� Des
photos de vol�es d'oiseaux ont �t� dispos�es sur un syst�me de coordonn�es
x-y. La localisation et l'envergure maximale des ailes des oiseaux ont �t�
d�termin�es. On a pr�sum� que les oiseaux volaient au m�me niveau d'altitude.
La longueur et la direction du vecteur total (sv) ainsi que la position de
la ligne t�te- t�te (kk) ont �t� calcul�es pour la formation enti�re selon
l'hypoth�se avanc�e. Les r�sultats d obtenus pour 4 formations (sur un total
de 22 observ�es) sont pr�sent�s sur la figure 20 a-d.
����������� Les r�sultats de toutes les formations �tudi�es ont v�rifi�
l'hypoth�se. L'�cart moyen des positions des oiseaux par rapport � la
position id�ale, selon l'hypoth�se �tait 5,8� d'angle. Les oiseaux volant
� l'ext�rieur de la stricte ligne de formation �taient en dehors de la
tendance g�n�rale des calculs avec un �cart de 50-70 % de la position
id�ale (fig. 20
a-d).
Electro-orientation et �lectronavigation
����������� A
partir d'un point de vue physique, les animaux en vol repr�sentent une
charge en mouvement, c'est-�-dire un courant �lectrique (courant de convection).
Pour autant qu'ils ne volent pas parall�lement aux lignes de champ magn�tique
terrestre, ils sont sujets � une force qui est, parmi d'autres, proportionnelle
� la vitesse de vol (force de Lorentz). Cette force donne lieu � son tour
� un potentiel �lectrique (effet Hall). En m�me temps un champ magn�tique
additionnel est produit autour des animaux en vol. Toutes ces grandeurs
physiques, qui sont indubitablement pr�sentes, sont en si minime proportion
que leur perception devrait sembler improbable. D'autre part, les biologistes
sont r�p�titivement surpris des possibilit�s qui existent dans le monde
animal. Des informations dont l'intensit� de puissance approche le niveau
du bruit de fond thermique, voire est noy�e dedans peuvent n�anmoins �tre
toujours per�ues par divers groupes d'animaux. C'est pourquoi, si les
charges �lectrostatiques des animaux en vol interviennent dans une interaction
complexe avec le champ magn�tique terrestre, et si les nuances r�sultantes
sont utilisables par l'animal, l'orientation du champ magn�tique des oiseaux
et des insectes pourrait �tre expliqu�e rationnellement. Une perception
magn�tique sp�ciale serait donc de ce fait non n�cessaire.
 |
�����
����������� Il est bien connu que les pigeons rentrant au pigeonnier ainsi que d'autres oiseaux entament un vol en spirale avant de prendre une direction particuli�re. Un tel comportement serait significatif si les animaux �taient capables de contr�ler les relations de forces
����������� a) qui
son produites � chaque angle de position entre le corps propre de l'individu
en mouvement et le champ magn�tique,
����������� b) qui se modifient avec la vitesse angulaire ou le rayon
du cercle parcouru,
����������� c) qui change avec l'altitude, selon le trac� des lignes de
potentiel et la charge �lectrostatique du corps.
Moins hypoth�tique est une facult� de navigation d�tect�e chez les sauterelles,
laquelle est bas�e sur la valeur de charge [36].
Si, durant un vol d�fini, non entrav�, dans le courant laminaire d'un
tunnel a�rodynamique, une sauterelle est entour�e d'une atmosph�re d'ions
n�gatifs (� des concentrations qui peuvent �galement se retrouver dans
la nature en certaines circonstances : 1,2.104 ions/cm�), elle
va r�agir imm�diatement par une diminution de la prise de nourriture (la
perte moyenne de puissance d�pendant de l'�ge: 300 mp � 50
mp) et une augmentation concurrente de la force ascensionnelle (fig.
21). Le valeur de seuil de concentration est d'environ 1.103
petits ions n�gatifs. La raison de la modification de la force nourriture/ascension
est une modification de la fr�quence des battements d'ailes qui correspond
� un effet "on-off" (effet ouvert-ferm� comme celui d'un interrupteur).
Apr�s une addition d'ions n�gatifs, la fr�quence se r�duit � 3 battements
par seconde. En m�me temps, une phase de neutralisation� entre les ailes
avant et les ailes arri�re augmente. Elle est probablement contr�l�e par
le syst�me nerveux central et peut �tre suivie au moyen d'un cin�matographe
� haute fr�quence et d'un stroboscope.
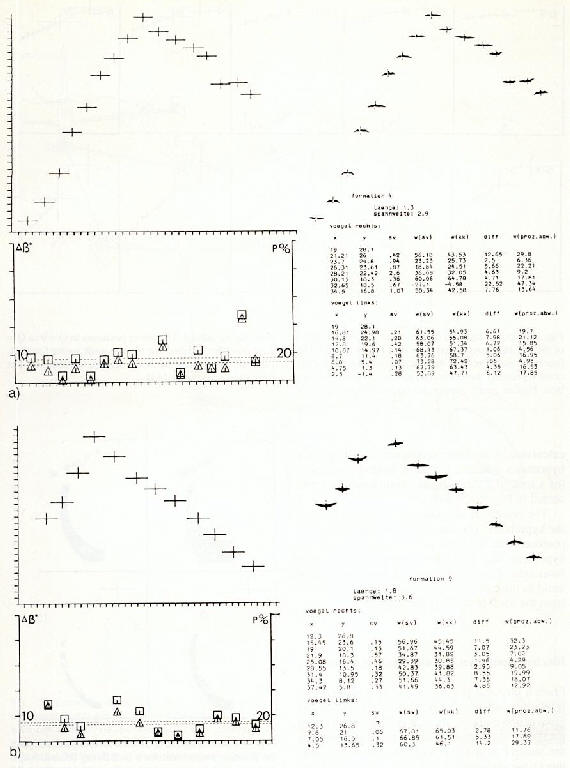 |
Fig. 20 a-b (voir l�gende globale sous fig 20 c-d)

�����������
Fig. 20 a-d: Etude de formations d'oiseaux
en vol en vue de v�rifier l'hypoth�se. La longueur du corps et l'envergure
des ailes (entre parenth�ses) respectivement pour a, b, c, d: 1,3 (2,9);
1,8 (3,6); 1,4 (2,6). Db
(triangles), angle entre le vecteur r�sultant et la ligne t�te-t�te (w(sv)
� w(kk)) P% (carr�s), Db comme pourcentage de l'angle inclus (voir fig. 16c).
Pour chaque formation, les groupes de donn�es sup�rieures et inf�rieures
se r�f�rent respectivement � des oiseaux volant � gauche et � droite de
l'oiseau de t�te [43].
Une augmentation
de la concentration de la charge spatiale dans l'environnement de l'animal
en vol repr�sente en fait une modification de sa charge statique. Comme
cela a �t� d�montr� lors d'exp�riences, cette modification est en corr�lation
avec l'altitude du vol.
����������� On pourrait conclure � partir de ce r�sultat, que les sauterelles
qui entreprennent elles aussi des migrations en groupes, sont capables
de naviguer en fonction des densit�s en ions. La densit� en ions est �
son tour en �troite corr�lation avec le niveau de potentiel du champ �lectrique
naturel. Des d�ductions similaires devraient �galement �tre appliqu�es
aux migrations d'oiseaux. Si ces oiseaux sont capables de percevoir indirectement
des niveaux de charge statique, il est aussi possible que leur navigation
soit bas�e sur ces valeurs.�
 |
����������� Plus petit
est l'insecte, plus la modification de la force ascensionnelle est perceptible
dans le champ �lectrique atmosph�rique vertical, tout simplement � cause des
composantes des forces de Coulomb. Selon la direction du champ, l'animal en
vol sera soit pouss� passivement vers le haut ou vers le bas. Cet effet est
maximal parmi les plus petites esp�ces d'insectes. Dans le champ d'un condensateur
de moins 5000 V/m, les animaux, se comportant comme un di�lectrique, peuvent
�tre graduellement "aspir�s vers l'ext�rieur". Dans le cas d'insectes
tr�s velus comme les bourdons terrestres (Bombus terrestris), avec
une tension (voltage) �lectrostatique totale d'environ 100 Volts (40 % d'humidit�
relative), une composante de force d'environ 2 % de la masse du corps est
engendr�e dans un m�me champ.
����������� Les vols d'hirondelles en altitudes �lev�es ou basses d�pendent
des conditions m�t�orologiques. Ceci est fonction de leurs proies, les insectes,
lesquels volent � diff�rentes hauteurs au cours des jours. Ceci pourrait s'expliquer
de mani�re rationnelle par les variations des forces ascensionnelles mises
en �vidence lors des essais en laboratoire. Ces forces ascensionnelles sont
d�termin�es par les charges �lectriques individuelles de chaque insecte et
par les valeurs de champ �lectrique atmosph�rique d�pendant des conditions
m�t�orologiques.
����������� Il y a encore un autre ph�nom�ne physique dans lequel des
informations peuvent �tre dissimul�es. Des abeilles volant par beau temps
non perturb�, par exemple, sont charg�es jusqu'� une certaine valeur durant
leur vol, depuis la ruche jusqu'� la source de nourriture. Cette valeur
atteint un �tat d'�quilibre d�pendant d'une dur�e d�finie. Lorsque l'abeille
atterrit sur une fleur, la plupart des charges accumul�es seront drain�es
de son corps (voir aussi fig.
3 et fig.12b).
Une nouvelle charge de m�me grandeur pourrait �tre accumul�e durant le
vol de retour vers la ruche. Si les abeilles sont capables de percevoir
un sch�ma spatio-temporel des charges �lectriques, ceci semblerait �tre
une possibilit� de transmission d'informations concernant les distances.
����������� En supposant que la charge est d'une importance fondamentale
pour les oiseaux et les insectes lors de leur navigation et dans leur
orientation, la d�charge devrait �tre associ�e avec la perte d'orientation
et la "nervosit�". Des calculs et des exp�rimentations impliquant
la charge d'insectes au moyen de facteurs �lectriques atmosph�riques naturels
montrent que les animaux sont d�charg�s, soit dans l'air, soit au sol,
bien avant qu'un orage �clate. Les abeilles pr�sentent � l'�vidence une
irritabilit� accrue durant cette p�riode. Ceci indique une connexion possible
entre la d�charge �lectrostatique, la perte d'orientation et l'irritabilit�
[37].
M�canismes de perception des champs �lectriques � extr�mement basses fr�quences (de 1 Hz � 3 kHz) et � tr�s basses fr�quences (de 3 kHz � 30 kHz).
����������� En
consid�rant l'influence des champs �lectriques sur l'homme et sur les diff�rentes
esp�ces animales, il y a plusieurs m�canismes ind�pendants qui peuvent avoir
des effets physiologiques primaires:
����������� a) les courants de d�placement;
����������� b) la diathermie;
����������� c) l'�lectrophor�se et l'�lectro-osmose;
����������� d) l'accumulation de charges locales caus�es par la r�sistivit�
(conductivit�) et l'effet semi-conducteur des tissus;
����������� e) les microvibrations, les pi�zo-r�sonances, le contr�le des
m�cano-r�cepteurs;
����������� f) les effets Josephson sur les membranes, les principes de coh�rence
avec la r�sonance dans les plages de dispersion alpha, les lasers � semi-conducteurs.
Si l'on consid�re la fr�quence et l'amplitude de l'intensit� du champ impliqu�, certains postes de la liste qui pr�c�de sont pr�dominants. Il faut avoir pr�sent � l'esprit qu'une plus forte intensit� de champ n'aura pas automatiquement une influence plus puissante. Au contraire, des exp�rimentations d�j� r�alis�es, montrent que l'�nergie est transf�r�e s�lectivement par diff�rents canaux selon la fr�quence et l'amplitude et que chaque canal r�pond � une largeur de bande particuli�re de fr�quences. Plus faible est l'apport d'�nergie, plus �lev�e est la probabilit� d'existence d'une fonction d'information associ�e, plus la plage de champ doit �tre observ�e de mani�re sp�cifique et circonspecte. Les r�actions aux champs extr�mement puissants et inhabituels sont comparables � l'effet d'un facteur de stress non sp�cifique. Lorsque ceci se produit, des courants de d�placement relativement puissants peuvent appara�tre et �tre enregistr�s dans l'organisme (souris: n = 10 kHz si E = 10 kV/m; I induite = 10-6 A). Il peut de ce fait na�tre des tr�s fortes densit�s de courant dans certaines voies particuli�rement conductrices. L'existence de� processus d'�lectrophor�se ou d'�lectro-osmose ne peut �tre �cart�e dans ce cas. Les m�mes effets devraient �tre engendr�es par une accumulation locale de charges.
Microvibrations
�����������
Avec des intensit�s de champ �lectrique de 5 kV/m, il a �t� possible
gr�ce � un dispositif d'interf�rence laser et d'une photodiode, de d�tecter
in vitro les microvibrations d'une cuticule d'insecte et des vibrisses
(moustaches) d'une souris (fig. 22). Diff�rentes parties de la cuticule
de l'insecte ont �t� �quip�es d'une petite bande de miroir de 2 mm d'�paisseur
pour r�fl�chir le rayon laser, tandis que les mouvements des vibrisses
de la souris ont �t� enregistr�es comme des modifications de l'intensit�
du rayon laser. Les oscillations m�caniques d�clench�es dans le corps
de l'insecte �taient soit transmises � l'air environnant sous la forme
d'ondes sonores, soit transmises � de l'eau environnante et ainsi photographi�es
(fig. 22b)
[41].
Lorsque le champ �lectrique atteint 10 kV/m, les ondes sonores peuvent
�tre facilement entendues gr�ce � un st�thoscope ou � un micro directionnel.
D'autres effets comme la pi�zo-�lectricit�, l'�lectro-contraction et les
interactions entre le champ et les charges statiques r�manentes, peuvent
�tre impliqu�s dans les r�sultats ci-dessus.
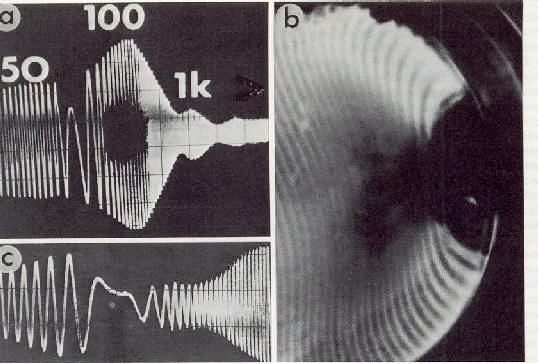 |
�����������
Fig. 22: Microvibrations sur la surface
du corps �d'insectes et d'autres animaux dans un champ �lectrique.
��������������� a) Corps d'insecte: oscillogramme obtenu
� partir d'un dispositif � interf�rence laser.
��������������� b) Corps
d'insecte: photo d'ondes sonores m�caniques transmises � de l'eau. L'enregistrement
qualitatif lorsque E = 5 kV/m; l'amplitude de vibration d�pend de la r�sonance
structurelle (ici, environ 100 Hz).
��������������� c) Vibration
d'une vibrisse de souris in vitro, dans un champ �lectrique de 30 kV/m
de fr�quences variant dans le temps.
Effets de pointes
����������� Une
question se pose quant � savoir si les animaux ont des aires localement
d�finies de t�guments qui absorberaient des charges de mani�re pr�dominante
et qui, comme des paratonnerres et des �metteurs de feux Saint Elme auraient
un effet de pointe et ainsi des puissances de champ augment�es. Ces aires
et ces structures pr�senteraient alors des amplitudes de vibrations particuli�rement
�lev�es en champs alternatifs ou bien rendraient possible une stimulation
m�canique ad�quate m�me avec de faibles intensit�s de champ.
����������� En vue d'approcher ce probl�me, un nuage d'a�rosol de CsCl
(chlorure de caesium) en solution aqueuse a �t� pulv�ris� � 50 cm devant
l'entr�e d'une ruche. Les particules lourdes d'a�rosol se sont d�pos�es
relativement vite alors que les plus l�g�res son soit rest�es en suspension,
soit ont �t� absorb�es par les abeilles en vol, selon l'intensit� de champ
�lectrique et la convection. Les zones d'absorption augment�e d'a�rosol
pouvaient alors �tre mesur�es par micro-analyse � diffraction de rayons
X (fig. 24).
La pr�sence de caesium �tait particuli�rement �vidente dans les aires
� pilosit� �lev�e. En vue d'obtenir des informations plus d�taill�es,
au sujet des micro-zones, une aile d'abeille a �t� utilis�e comme mod�le
de fonction dans l'exp�rience. La figure 23 montre la morphologie de l'aile.
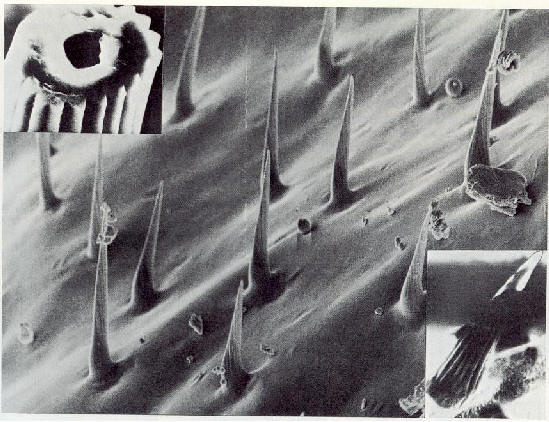 |
Fig. 23: Topographie d'une aile d'abeille. Les excroissances sont creuses (au dessus � gauche) et remplies d'�lectrolyte. In effet �lectrique de pointe peut �tre atteint sur l'excroissance sous l'influence d'un rayonnement �lectromagn�tique (en dessous � droite). Mod�le fonctionnel de l'effet de succion de pointes charg�es sur les charges de l'espace environnant.
�������
Les
excroissances �pineuses sont r�guli�rement espac�es sur une surface lisse.
Ces excroissances sont creuses et remplies d'un �lectrolyte (voir
figure 23). Sous l'influence d'un champ �lectromagn�tique,
un effet de pointe a �t� observ� sur les excroissances lorsqu'elles sont
statistiquement charg�es (section agrandie en dessous � droite). Cependant,
les conditions �taient non physiologiques. L'aile a ensuite �t� coup�e
� la base et de petits fils ont �t� fix�s aux sections des veines au moyen
d'une laque conductrice au graphite. Ceci a permis d'amener le syst�me
d'�lectrolyte, et de l�, les pointes des ailes � des niveaux diff�rents
de potentiels et de polarit� en les comparant � la terre. L'effet de pointe
a �t� test� en amenant le mod�le d'aile en contact avec un a�rosol de
chlorure de caesium qui �tait soit neutre, soit charg� � un potentiel
de 20 kV, positif ou n�gatif.
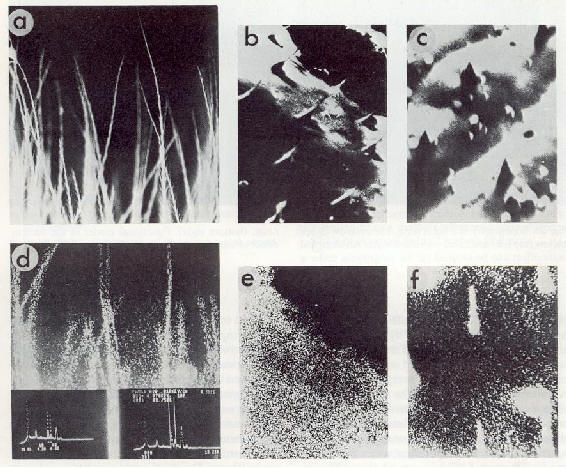 |
��������
Fig. 24: D�monstration de l'adh�rence de charge sur la couche externe du corps
d'une abeille et de l'aptitude qu'ont les parties expos�es du corps �
accumuler les charges lorsqu'elles sont mises en contact avec un a�rosol
de charge oppos�e.
a)
image en microspectroscopie �lectronique de poils thoraciques;
b)
image en microspectroscopie �lectronique d'une section d'aile. Aile �lectriquement
neutre pendant l'exp�rience;
c)
image de dispersion d'une section d'aile. Le syst�me d'�lectrolyte de
l'aile sur potentiel durant l'exp�rience.
d)-
f) images correspondantes de la distribution de l'�l�ment au moyen de
spectroscopie du Cs aux rayons X.
e)
distribution du Cs sans diff�rentiation;
f)
distribution pr�pond�rante du Cs sur les pointes avec la canalisation
d'�lectrolyte.
La
preuve pr�cise a �t� obtenue par trac� de la distribution des forces de
Coulomb au moyen d'un microspectroscope. Les r�sultats ont montr� une
stimulation m�canique de ces structures dans le champ �lectrique alternatif
et ont ainsi r�v�l� une forme de s�lectivit� attribuable � un ph�nom�ne
de r�sonance. Ceci implique que l'assimilation des champs �lectriques
est finalement r�duite � l'assimilation d'une �nergie m�canique. D'autre
part, nous savons que les animaux plac�s dans des conditions normales,
r�agissent � ces faibles �nergies �lectriques et magn�tiques de mani�re
telle qu'une transduction en �nergie m�canique physiologique semblerait
devoir �tre �cart�e. Des m�canismes d'amplification sont n�cessaires ici,
avec des d�tecteurs de m�canismes capables d'avancer de pair avec les
d�couvertes de la physique contemporaine.
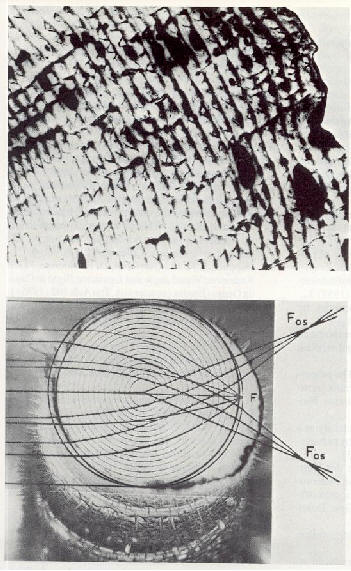 |
Avec nos remerciements au Prof. Dr. E. Morgenstern, Institut de Microscopie �lectronique, Universit� de la Sarre.���
Cette
contribution concerne seulement la transmission d'informations gr�ce �
des champs �lectriques statiques et alternatifs � basse fr�quence. Nous
travaillons parall�lement sur la distribution de l'information au moyen
de champs magn�tiques et d'oscillations �lectromagn�tiques. Une pr�sentation
des relations connues aujourd'hui requi�rent un article s�par�. Les m�canismes
organiques concernant ce type de transmission d'�nergie doivent �galement
�tre pr�sents chez l'animal et il est possible qu'on y d�couvre une fonction
d'amplification ou la focalisation sur une largeur de bande particuli�re
des signaux �lectromagn�tiques. Les antennes d'une gu�pe fournissent une
bonne illustration (fig.
25). Le segment montre une coupe de la paroi de l'antenne dans
la forme de l'image d'une transmission �lectronique.
�������� On peut apercevoir le milieu stratifi� coaxialement et non homog�ne,
qui repr�sente pour les ondes �lectromagn�tiques tombant longitudinalement
une lentille cylindrique - peut-�tre � double foyer -. Une condition pr�alable
est que l'�paisseur des couches concentriques doit �tre faible, mais la dimension
externe plus grande, comparativement � la longueur d'onde. Un hypoth�tique
d�tecteur de micro-ondes plac� � l'extr�mit� ou � l'ext�rieur de la lentille-antenne
pourrait ainsi capter une plus forte intensit� que celle des ondes re�ues.
L'intensit� est alors une fonction de l'angle d'azimut � l'endroit du r�cepteur.
Les investigations d�taill�es concernant la transmission d'informations gr�ce
� des champs �lectriques et magn�tiques et gr�ce � des oscillations �lectromagn�tiques
ont jusqu'ici �t� n�glig�es. Plusieurs parmi nos raisonnements bien qu'hypoth�tiques
� l'origine se sont r�v�l�s valides lorsqu'ils ont �t� confront�s aux exp�rimentations.
Le principe d'exploitation des formes pr�sentes d'�nergie pour la communication
et l'orientation existe donc bien dans les organismes vivants, non seulement
dans l'eau, mais aussi sur terre, gr�ce � la mise � profit des structures
du corps et des mat�riaux.
(1)
Note du traducteur: Un vieux dicton nous dit "Lorsque
le chat en faisant sa toilette, d�passe l'oreille avec la patte, c'est qu'il
va pleuvoir". L'accroissement de l'humidit� ambiante pr�c�dant la pluie
n'incite t-elle pas le chat � r�tablir avec des composants de sa salive, les
potentiels de certains poils sensitifs situ�s autour et dans l'oreille?
(2)
Un �lectret est un corps �lectris� de mani�re permanente,
dont les mol�cules ont pris une orientation donn�s sous l'effet d'un champ
�lectrique temporaire (certains micrphones fonctionnent selon ce principe).
R�f�rences bibliographiques
[1]
Altmann G., Warnke U. : "Einfluss unipolar geladener Luftionen
auf die motorische Aktivit�t der Honinbienen", Apidologie, 2
pp. 309-317 (1971).
[2] Altmann G., Warnke
U. : "Unspezifische analoge Reaktion von M�usen auf starke elektrische
Feldrer und Elektrische Abschirmung"; Z. Physik. Med.,
2, pp. 137-143 (1978).
[3] Athenstaedt
H. "Pyroelectric and piezoelectric properties of vertebrates",
Ann. New York Acad. Sci., 238 pp. 68-94 (1974).
[4]
Basett C.A.L. "The biological significance of piezoelectricity",
Calcif. Tiss. Res. 1 p. 252 (1968).
[5]
Becker L. "Untersuchungen �ber das Heimfindeverm�gen der Bienen"
Z. vergl. Physiol., 41 p.1 (1958).
[6]
Becker G. "Communication between termites by biofields"
Biol. Cybernetics, 26 pp. 41-44 (1977).
[7]
Bergh J.E. "Insect take-off activity as a response to meteorological
stimuli effective over long distances" Th�se de Doctorat, Uppsala
(1985).
[8]
Bigu-Del-Blanco J., Romero-Sierra C. "The properties of bird feathers
as concerse piezoelectric transducers and as receptors of microwave radiation",
Biotememetry, 2 pp. 341-364 (1975).
[9]
Cope F.W.: "Piezoelectricity and pyroelectricity as a basis for
force and temperature detection by nerve receptors", Bull. Mathem.
Biol., 35, p.31 (1973).
[10]
Dornfeldt K. "Statistische Beziehungen der Wettflugsgeschwindigkeit
von Brieftauben mit meteorologischen, geophysikalischen und wettflugorganisatorischen
Faktoren": Verh. Dtsch. Zool. Ges., Erlangen, 334 (1977).
[11] Drost R., "Zugv�gel
perzipieren Ultrakurzwellen" Vogelwarte, 2 pp. 57-59 (1949).
[12]
Edwards D.K. "Effects of artificially produced atmospheric electrical
fields upon the activity of some adult diptera" Can. J. Zool.,
38 pp. 899-912 (1960).
[13]
Ellenby C., Smitz L. "Observations on the piezoelectric properties
of nematode cuticle" ; Comp. Biochem. Physiol.
26 pp. 359-363 (1968).
[14] Erickson
E.H. "Surface electric potentials on worker honey-bees leaving
and entering the hive" J. Apicult. Research, 14, pp. 141-147
(1975).
[15]
Eskov E.H., Sapozhnikov A.M.: "Generation and perception of electric
fields by Apis Mellifera"; Zool. Z.: 53, 5, pp. 800-802 (1974).
[16] Eskov
E.K., Sapozhnikov A.M.: "Mechanisms of
generation and perception of electric fields by honeybees"; Biophysik,
21, 6 pp. 1097-1102 (1976).
[17] Exner
S. "�ber die elektrischen Eigenschaften der Haare und Federn"
Pfl�gers Arch. 61/63 pp. 427-449 (1895).
[18]
Fukada E. "Mechanical deformation and electrical polarization
in biological substances" Biorheology, 5, pp. 199-208 (1968).
[19]
Galuszka H., Lisiecky J. "Certain reactions in honeybees to the
flow of electric current of different parameters" Zoologica Poloniae;
19, 2, pp.197-211 (1969).
[20]
Gemmel U. "Histologische Befunde an den Nebennierenrinde von Ratten
und M�usen nach Einwirkung elektrischer Felder und bei elektrischer Abschirmung"
Th�se, Saarbr�cken (1978).
[21]
Gottschaldt K.M. "Mechanoreceptors in the beaks of birds"
Rhein, - Westfal. Akad. d. Wiss., (Symposium Mechano-reception) 53, pp.
109-113 (1976)
[22]
Heppner H.F., Haffner J., "Electromagnetic fields for communication
in birds flocks"; A model symposium and workshop on the effects
of low-frequency magnetic and electric fields on biological communication
processes at the 6th annual meeting of the Neuroelectric Society,
Colorado, Vol. 6 (1973).
[23]
Heppner H.F., Convissar J.L., Moonan D.E., Anderson J.G.T. "Visual
angle and formation flight in Canada Geese (Branta Canadensis)";
The Auk, 102, 1 (1985).
[24]
Heuschmann O. "�ber die elektrischen Eigenschaften der Insektenhaare"
Z. vergl. Physiol., 10, pp. 594-664 (1929).
[25] H�sing J.O., Stawory
P. "Zur Frage der Ausl�sung des Jungbieneneinfluges";
Wiss. Z. Martin Luther Univ. Halle-Wittemberg, Math.-Nat. 8/6 pp. 1121-1122
(1959).
[26] Hummel D., "Die
Leistungserparnis beim Verbandsflug", J. Orn., 114, pp. 259-282
(1973).
[27]
Iggo A., Gottschaldt K.M. "Cutaneous mechanoreceptors in simple
and in complex sensory structures"; Rhein.-Westf.. Akad.
d. Wiss. 53 Symposium Mechanoreception, pp. 153-174 (1976).
[28] Jaeger
P. "Raumladungmessungen im Bienenwachs w�hrend der Erstarrung und im festen
Zustand" Anal. Phys. 5, 21 pp.
481-502 (1934).
[29]
Maw M.G. "Behaviour of an insect on an electricallly charged surface"
Canad. Entomol. 39, pp. 391-393 (1961)
[30]Nachtigall W. "Phasenbeziehungen
der Fl�gelschl�ge von G�nsen w�hrend des Verbandfl�ge in Keilformation";
Z. vergl. Physiol. , 67 pp. 414-422 (1970).
[31]Necker R. "Temperature
sensitivity of slowly-adapting mechanoreceptors on the beaks of pigeons";
Rhein.-Westf.. Akad. d. Wiss. 53 Symposium Mechanoreception,
pp. 115-121 (1976).
[32] Poncy R. "
Formations angulaires des vols de quelques esp�ces d'oiseaux migrateurs
et les r�actions observ�es" Ornitholog. Beobachter,
38, pp. 18-19 (1941).
[33] Schua L. "
Untersuchungen �ber den Einfluss meteorologischer Elemente auf das
Verhalten der Honigbienen (Apis mellifica)"; Z. vergl. Physiol.,
34 pp. 258-277 (1952).
[34] Straubel H. "Experimente
mit makroskopischen geladenen Teilchen"; Physikal. Bl�tter, 28, pp. 65-70 (1972).
[35]
Tshernyshev W.B., Ershova N.J., Tikhonova E.V., Shakanova E.M. "Influence
of electrical charges of earth surface on some soil arthropoda";
Pedobiologia, 13, pp. 437-440 (1973).
[36]
Warnke U. "Untersuchungen des Einflusses unipolarer atmosph�rischer
Ionen auf die motorische Aktivit�t von Insekten" Staatsexamensarbeit
f�r Lehramt an H�heren Schulen 1971 (non publi�).
[37] Warnke U. "Physikalisch-physiologische
Grundlagen zur luftelektrisch bedingten "Wetterf�hlichkeit"
der Honigbiene (Apis Mellifica)". Diss., Saarbr�cken (1973).
[38] Warnke U. "Insekten
und V�gel erzeugen elektrische Felder." Umschau,
75, 15 p. 479 (1973).
[39]
Warnke U. "Effects of electric charges on honeybees"
Bee Res. Ass. 57, 2, pp.50-56 (1976).
[40]
Warnke U. "Die Wirkung von Hochspannungswechselfeldern auf das
Verhalten von Bienensoziet�ten", Z. angew. Entomol.,
82 1, p.88 (1976).
[41]
Warnke U. "Bioelectric and biomagnetic qualities on skins of animals,
influenced by meteorological facts"; II Bioclimatic results of
static and electromagnetic facts. TU Munich (1976).
[42]
Warnke U. "Die elektrostatische Aufladung und Polarisierbarkeit von Insektenintegumenten";
Verh. Dtsch. Zool. Ges., Erlangen, p332 (1977)
[43]
Warnke U., "Avian flight formation with the aid of electromagnetic
forces: a new theory for the formation alignment of migrating birds
", J. Bioelectricuty, 3, 3 pp. 493-508 (1984).
[44]
Warnke U., "Relevanz elektrischer Biofelder" ; Umschau
6, pp.336-343 (1986)
[45]
Warnke U., "Thermography of honeybee colonies in winter influenced
by high voltage electric fields"; Z. Entomol. 104,
1, pp. 69-73 (1987).